|
MUSCLE SYSTEM
GLR CELLS
 Click pictures for new window with figure and legend, click again for high resolution image Click pictures for new window with figure and legend, click again for high resolution image
1 Overview
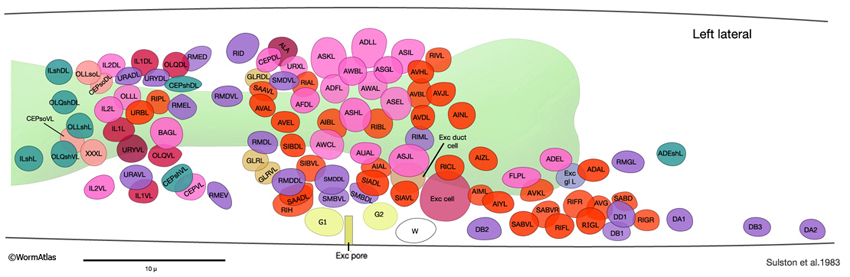
Three pairs of GLR cells (GLRDL/R, GLRL/R, GLRVL/R) lie within the pseudocoelom at the level of the pharyngeal isthmus, slightly posterior to the nerve ring (GlrFIG 1). They are arranged in a sixfold symmetrical manner (GlrFIG 2). The cell bodies of the GLRs lie near the dorsal and ventral insertions of the muscle arms of the head, where the muscle arms enlarge to dive toward the isthmus of the pharynx. This close physical proximity may reflect the common lineage of the GLRs and the body wall muscles because both derive from the MS lineage. Their sister cells also include the head mesodermal cell, the pharyngeal musculature, and the coelomocytes (For more information see sections for: Head Mesomdermal Cell, Nonstriated Muscle, Coelomocytes).
GlrFIG 1: Nuclei positions in the nerve ring. Based on Sulston et al., 1983.
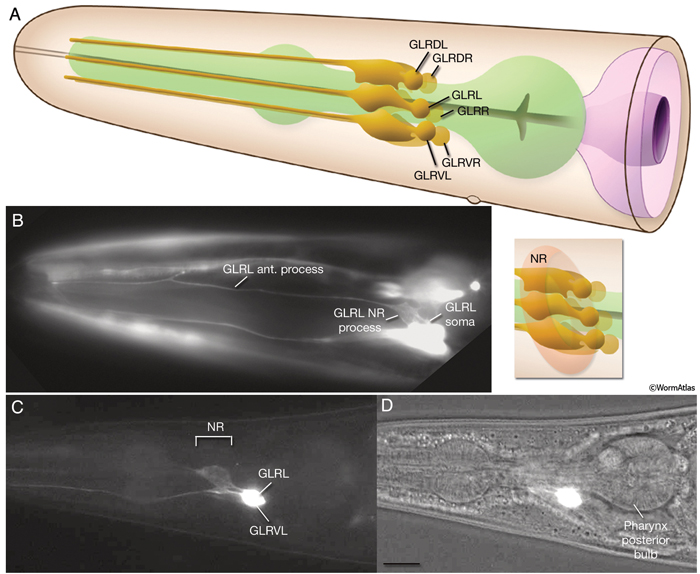
GlrFIG 2: GLR cells and their processes. A. Cell bodies of GLR cells are aligned dorsoventrally, just posterior to the nerve ring on each side of the pharynx isthmus. A thin, leaf-like process grows out of each cell to wrap the interior of the muscle-neuron plates of the nerve ring (inset). At the anterior edge of the nerve ring, this process narrows down to a thin rod that joins the labial process bundles and travels anteriorly to the level of the anterior end of the pharynx, where it peters out. B. Epifluorescent image from a transgenic animal expressing the gly-18::GFP reporter gene. The GLRL soma is seen on the lateral side of the isthmus. Anteriorly to the nerve ring region, the GLR process becomes quite thin and continues to travel along the lateral side of the pharynx, left lateral view. (Strain source: C.E. Warren, A. Krizus, and J.W. Dennis.) C. Epifluorescent image from a transgenic animal expressing the C05D9.1::GFP reporter gene, left lateral view. The anterior GLR processes flatten out at the nerve ring (NR) region. (Image source: R. Newbury. The Genome BC C. elegans gene expression consortium [McKay et al., 2004].) D. DIC image of the same animal as in C. Bar, 10 μm.
All six GLR cell somata lie posteriorly to the nerve ring, and their positioning reflects the tilt of the nerve ring. The pronounced medial turn of each head muscle arm occurs in close apposition to GLR cell bodies. Anteriorly, each GLR cell extends a thin, sheet-like process (GlrFIG 2B). On the outside, these processes are surrounded by the muscle plate that lies inside the motor end plate layer of the nerve ring, where muscles receive synaptic input from the nerve ring motor neurons (GlrFIG 2 and GlrFIG 3). Each of these processes wraps around roughly one third of the circumference of the isthmus, touching its neighbor on each side (GlrFIG 4, GlrFIG 5 and GlrFIG 3H). There is no direct connection of the GLRs to the pharyngeal basement membrane, which lies between the GLR cells and the pharynx, and there is no basement membrane separating the muscle arms from the GLR processes (White et al., 1986). Arms from each of the eight longitudinal rows of the head muscles run along specific GLR cells such that each GLRDL/R and GLRVL/R is associated with muscle arms from a single row and each GLRL and GLRR is associated with muscle arms from two rows. Gap junctions exist between GLR cells and the muscle arms and between GLRs and RME motor neurons (see Gap Junctions). However, GLR cells do not make gap junctions to one another nor are they involved in any chemical synapses.
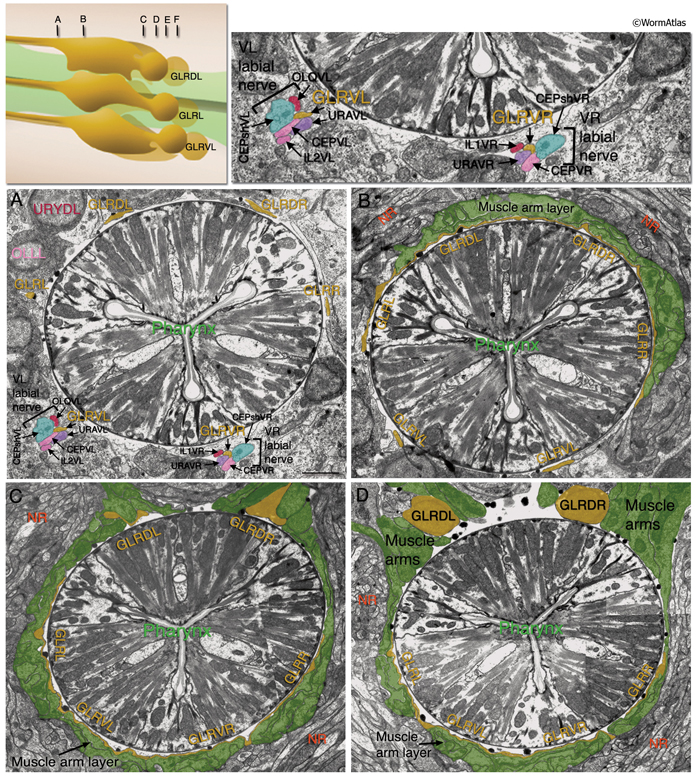
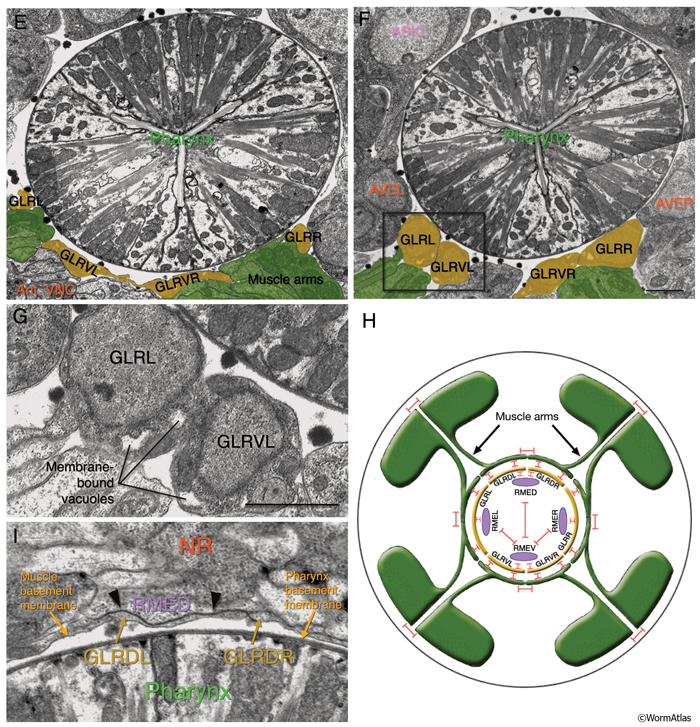
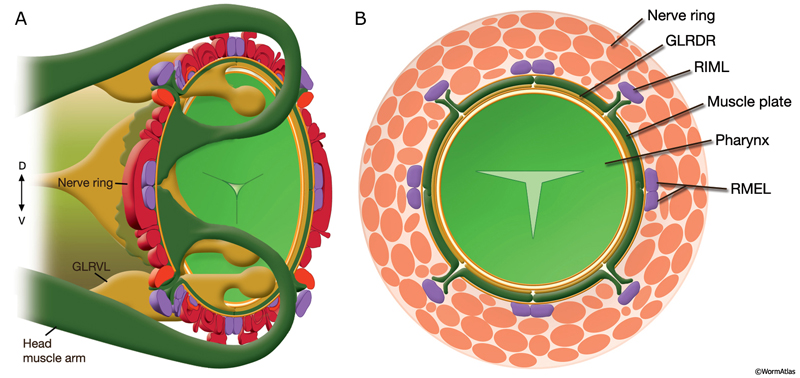
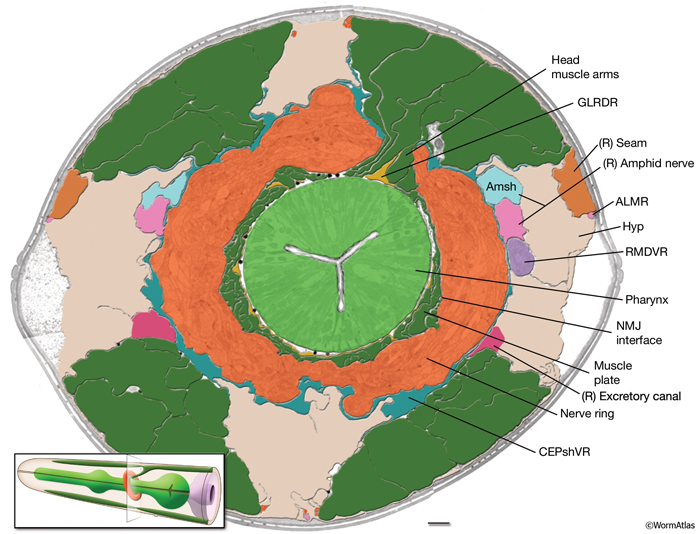 Anterior to the nerve ring, GLR processes narrow down into thin processes and continue anteriorly to peter out at the level of the junction of the pharynx and the buccal cavity without any terminal specializations (GlrFIG 2 and GlrFIG 3). Throughout their length, the anterior GLR processes run in the inner labial bundles, closely apposed to the IL1 dendrites.
The cytoplasm of the GLR cell body is electron dense and contains a distinctive collection of membrane-bound vacuoles. With TEM, these vacuoles formally look very similar to inclusions in the distal tip cells and the coelomocytes. This suggests an active endocytic or secretory function for GLRs.
2 Function of GLR Cells
Because of their location, connectivity pattern, and lineage, GLR cells are suggested to be mesodermal scaffolding cells that guide muscle arms to appropriate territories during development (White et al., 1986). The C. elegans myoD homolog hlh-1, which is expressed by lineal precursors of body wall muscle and is required for normal muscle function, is also expressed in GLR cells in late embryogenesis and larval stages. It has been suggested that hlh-1 might drive expression of cell-surface proteins that mediate recognition and contact between GLRs and head muscles (Krause et al., 1994). Similar to body wall muscle cells, GLR cells secrete type IV collagen, which is integrated into the basement membrane underlying the muscle (Graham et al., 1997; Norman and Moerman, 2000).
At the anterior end of the nerve ring, the sheet-like anterior processes of GLRs briefly seal the space between the end of the somatic basement membrane of the muscle and the basement membrane of the pharynx and the pseudocoelom. Hence, anterior to this region, the narrow space between the pharynx and outer tissues is designated as an accessory pseudocoelom (see Pericellular Structures) (Z. Altun and D.H. Hall, unpubl.). It is not yet clear if there is material exchange between these two spaces. When any one of the parental cells of the GLRs (MSaaaaaa for GLRDL/R; MSapaaaa for GRLL/VL; MSppaaaa for GLRR/VR) is killed in the embryo, the worms hatch late and arrest as starved L1-stage animals. In these animals, nerve rings are displaced anteriorly and there is widespread degeneration and vacuolation in neurons and hypodermis, which may result from disruption of the GLR seal (A. Chisholm, pers. comm.).
3 List of GLR Cells
GLRL; MSapaaaad
GLRR; MSppaaaad
GLRVL; MSapaaaav
GLRVR; MSppaaaav
GLRDL; MSaaaaaal
GLRDR; MSaaaaaar
|
4 References
Graham, P.L., Johnson, J.J., Wang, S., Sibley, M.H., Gupta, M.C. and Kramer, J.M. 1997. Type IV collagen is detectable in most, but not all, basement membranes of Caenorhabditis elegans and assembles on tissues that do not express it. J. Cell Biol. 137: 1171-1183. Article
Krause, M., White Harrison, S., Xu, S-Q., Chen, L. and Fire, A. 1994. Elements regulating cell- and stage-specific expression of the C. elegans MyoD family homolog hlh-1. Dev. Biol. 166: 133-148. Abstract
McKay, S.J., Johnsen, R., Khattra, J., Asano, J., Baillie, D.L., Chan, S., Dube, N., Fang, L., Goszczynski, B., Ha, E., Halfnight, E., Hollebakken, R., Huang, P., Hung, K., Jensen, V., Jones, S.J.M., Kai, H., Li, D., Mah, A., Marra, M., McGhee, J., Newbury, R., Pouzyrev, R., Riddle, D.L., Sonnhammer, E., Tian, H., Tu, D., Tyson, J.R., Vatcher, G., Warner, A., Wong, K., Zhao, Z. and Moerman, D.G. 2004. Gene expression profiling of cells, tissues and developmental stages of the nematode C. elegans. Cold Spring Harbor Symp. Quantit. Biol. 68: 159-69. Abstract
Norman, K.R. and Moerman, D.G. 2000. The let-268 locus of Caenorhabditis elegans encodes a procollagen lysyl hydroxylase that is essential for type IV collagen secretion. Dev. Biol. 227: 690-705. Article
Sulston, J.E., Schierenberg, E., White, J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64-119. Article
White, J.G., Southgate, E., Thomson, J.N. and Brenner, S. 1986. The structure of the nervous system of the nematode Caenorhabditis elegans. Phil. Trans. Roy. Soc. Lond. 314B: 1-340. Article

This chapter should be cited as: Altun, Z.F. and Hall, D.H. 2009. Muscle system, GLR cells. In WormAtlas. doi:10.3908/wormatlas.1.9
Edited for the web by Laura A. Herndon. Last revision: April 23, 2013 |

|
|