|
|
|
THE DAUER EPITHELIUM
 Click pictures for new window with figure and legend, click again for high resolution image Click pictures for new window with figure and legend, click again for high resolution image
1 Overview
During the transition from the L2 to dauer stage, the larval body is remodeled to produce a slimmer physique, mostly through a reduction in the volume of epidermal tissues. The epidermis includes the hypodermis, surrounding the body at the exterior surface, as well as the seam cells and XXX cells, which are more specialized in function. (See also Hermaphrodite Epithelial Sections on Hypodermis, Seam and XXX cells). As this change is taking place, the epidermal tissues produce a unique new cuticle to enclose the dauer body, providing added protection against external damage and dehydration. The rigid dauer cuticle and elaborate lateral alae may also increase traction for more efficient body motion (see Dauer Cuticle). The epidermal cells produce basement membrane components for structural support, serve as fat storage depots and secrete hormones. These functions are shared by epidermal cells in both dauer and nondauer stages, although the relative importance of these functions may be shifted in dauers to emphasize protection and storage over growth and development (Burnell et al., 2005). In addition, the epidermal XXX cells perform a signaling role, which aids in regulating dauer arrest and recovery (Gerisch and Antebi, 2004; Schaedle et al., 2012).
2 Anatomy
2.1 Hypodermis
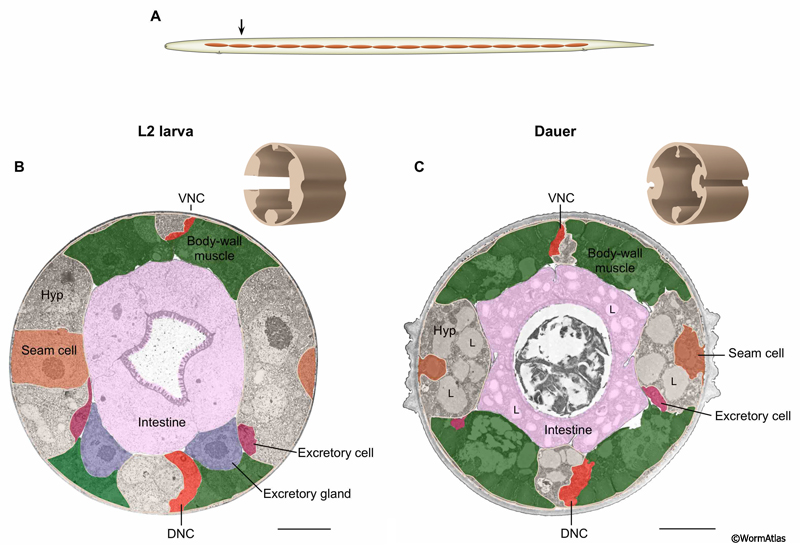
The body is surrounded by the hypodermis, which is comprised of four longitudinal ridges (right lateral, left lateral, dorsal and ventral) connected by cytoplasmic extensions to completely encircle all other tissues. The main hypodermis is a syncytium of individual hypodermal cells formed in the embryonic and larval stages. In dauers, the four hypodermal ridges shrink as a proportion of body size and volume through autophagy during the L2d stage (DEpiFIG 1) (Singh and Sulston, 1978; Meléndez et al., 2003). The hypodermis secretes a much thicker, reinforced body cuticle during the L2d to dauer transition, so that the dauer is far better protected against damage or dehydration (see Dauer Cuticle). Refractile bodies, possibly related to lipid storage droplets, have been described in the dauer hypodermis (DEpiFIG 3) (Vowels and Thomas, 1992).
DEpiFIG 1: Hypodermal tissue is shrunken and filled with lipid in dauer larvae. A. Illustration showing approximate position of sections in lower panels. B&C. Transverse sections from the anterior intestinal areas of L2 and dauer larva, respectively. 3D rendering of hypodermis from sections in brown shows footprint of tissue. Hypodermis (unshaded) occupies a smaller fraction of the body in the dauer than in the L2 with all four hypodermal quadrants strongly affected. In contrast, the body wall muscle appears roughly similar in size in L2 and dauer larvae (green, see Dauer Muscle section). In dauers, hypodermal shrinkage is due to autophagy (self digestion) (Meléndez et al., 2003). Note the presence of prominent lipid droplets (L) in the dauer hypodermis. VNC, ventral nerve cord; DNC, dorsal nerve cord; Hyp, hypodermis. Scale bar, 5 microns. (Image source: L2 [D. Riddle] N2 28-14 1627; dauer [D. Riddle] 50-2-1 1362.)
2.2 Seam Cells
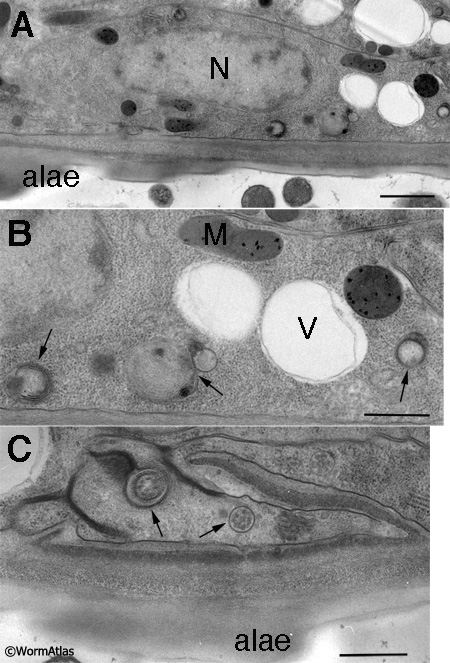
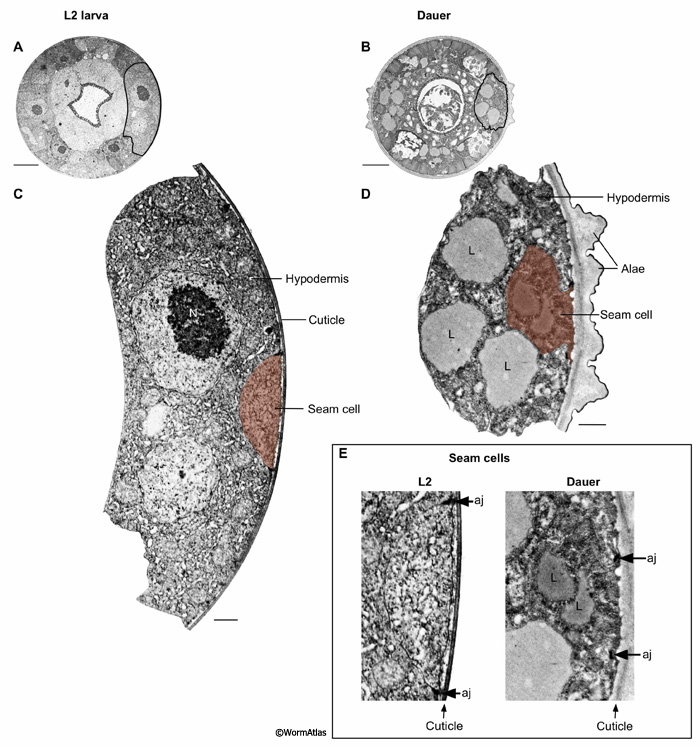
The seam cells are also profoundly shrunken in dauers due to autophagy during the L2d stage (DEpiFIG 2), and Golgi bodies are nearly absent (Singh and Sulston, 1978; Popham and Webster, 1979; Meléndez et al., 2003) (DEpiFIG 2). When L2d larvae molt into dauers, the seam cells produce characteristic dauer alae with a five-fold structure (see Dauer Cuticle) (Singh and Sulston, 1978). During the time of highest autophagy, seam cells (and hypodermis) feature many large seemingly empty vacuoles (DEpiFIG 2) in addition to many small autophagosomes. It is possible that these vacuoles may also play a role in cell shrinkage occurring in L2d and early dauer. They appear different (much less electron dense) in contents compared to the prominent lipid droplets that accumulate in the later dauer epidermis (DEpiFIG 1 and DEpiFIG 3).
DEpiFIG 2: Dauer seam cell showing enhanced autophagy. A. Low power micrograph of a dauer animal (daf-2(e1370) mutant). Here the seam cell is locally enlarged in size to accommodate the nucleus (N), and the cell lies directly apposed to the cuticular alae. The seam cell cytoplasm on the lower right side of the panel holds a cluster of interesting inclusions, shown at higher magnification in panel B. Autophagy is greatly enhanced in the seam cell during dauer formation (Meléndez et al., 2003). The seam cell and the neighboring hypodermis (top right corner of this panel) are also filled with large vacuoles at this stage. B. Arrows indicate several autophagosomes at various stages of processing. Each autophagosome is bordered by two concentric membranes, with small bits of cytoplasm enclosed within the inner membrane for degradation. C. Another section through the daf-2 seam cell at a position far away from the nucleus shows the seam greatly reduced in volume, but still enclosing several autophagosomes.
N, nucleus, M, mitochondrion, V, vacuole. Scale bar in A is 1 micron. Scale bars in B & C are each 0.5 micron. (Image source: Hall lab. A. F3, W390; B. F3, W391; C. H2, W426.)
|
3 Function
3.1 Lipid Storage
Lipid storage granules are prominent in the dauer hypodermis, emphasizing its role in nutrient storage (Burnell et al., 2005) (DEpiFIG 1 & DEpiFIG 3). The intestine also serves as a fat storage depot in dauers (DEpiFIG 1C). It is not known whether the nutrients stored in each tissue serve overlapping functions or exist to serve separate requirements.
3.2 Regulation of Dauer Formation
Genetic pathways acting within the hypodermal cells can have powerful effects on dauer tissue remodeling throughout the body. For example, a microRNA pathway that regulates cell fate decisions in the hypodermis has effects that radiate to other tissues (Liu and Ambros, 1989). In addition, a hormonal pathway operating in the hypodermis appears to control hormone levels throughout the body in response to environmental conditions (Antebi et al., 2000).
3.2.1 MicroRNA pathways
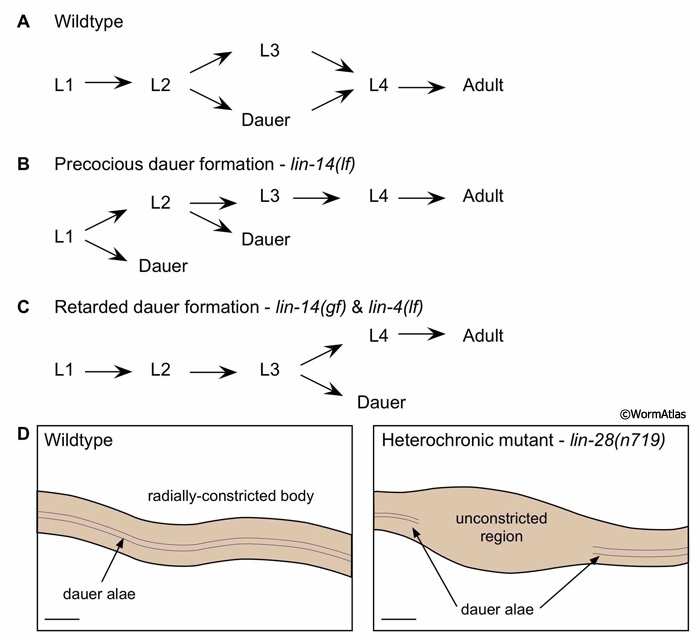
MicroRNA pathways coordinate many developmental transitions in C. elegans, including some aspects of dauer larval development. The microRNA, lin-4, controls dauer entry by negatively regulating expression of its target genes, lin-14 and lin-28, which affect hypodermal cell fate specification during development (Vella and Slack, 2005). Mutations in these genes can also affect dauer entry (Liu and Ambros, 1989). Some mutants, such as lin-14 loss-of-function (lf) alleles, form dauers precociously after the L1 stage rather than after the L2 (DEpiFIG 4). The lin-14(lf) precocious dauers possess most features of wildtype dauers, except that the gonads are smaller and more similar to those found in L2 larvae. By contrast, other mutants, such as lin-4(lf) and lin-14 gain-of-function (gf) mutants, delay dauer formation by allowing L3 larvae to molt into dauers. These dauers have enlarged reflexed gonads, typical of L4 larvae, but are otherwise normal-appearing dauers. In some cases, lin-4(lf) and lin-14(gf) animals are completely unable to form dauers. lin-28(lf) mutants and several of the lin-14 mutants show partial hypodermal remodeling. In these partial dauers, some body regions fail to radially constrict or produce dauer alae, while adjacent areas complete radial constriction and alae synthesis normally (DEpiFIG 4D).
3.2.2 Dafachronic acid pathway
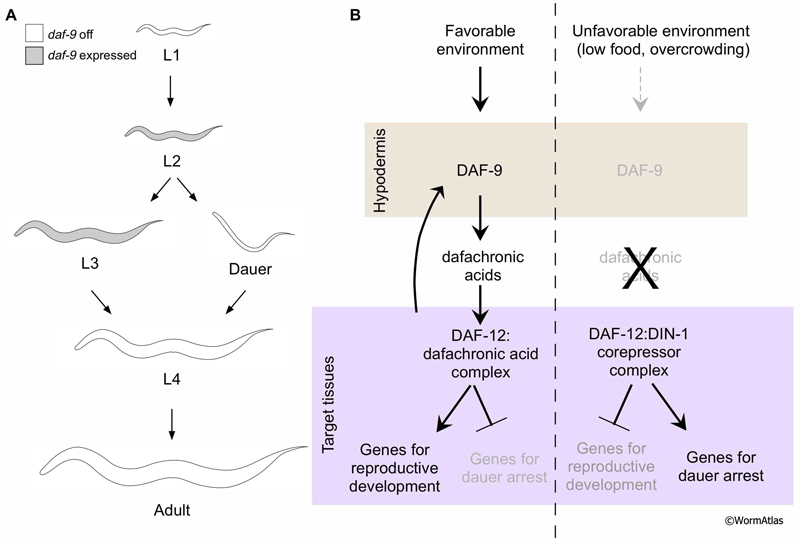
The DAF-12/nuclear hormone receptor (NHR) is a ligand-regulated transcription factor that performs dual roles in dauer formation and reproductive development (Antebi et al., 2000; Held et al., 2006; Motola et al., 2006). Favorable environmental conditions promote production of DAF-12 ligands, 3-keto-sterol derivatives named dafachronic acids (Motola et al., 2006). Liganded DAF-12/NHR directs the expression of genes needed for adoption of L3 fates, particularly in the hypodermal and gonadal cells (Antebi et al., 1998). Conversely, harsh environmental conditions reduce dafachronic acid synthesis, increasing the proportion of non-liganded DAF-12/NHR. In its non-liganded state, the DAF-12/NHR binds the DIN-1 corepressor, forming a complex which promotes expression of genes needed for dauer formation and may repress expression of genes for reproductive development (Ludewig et al., 2004).
Dafachronic acid is produced by a biosynthetic pathway modifying cholesterol. One of the major enzymes in this pathway is encoded by the gene, daf-9 (Gerisch et al., 2001; Jia et al., 2002; Mak and Ruvkun, 2004). An important site for daf-9 expression and function is the hypodermis, although daf-9 is expressed in several other tissues as well (Gerisch and Antebi, 2004, Mak and Ruvkun, 2004). Environmental factors influencing dauer arrest regulate hypodermal daf-9 expression and, consequently, dafachronic acid levels. Hypodermal daf-9 expression is observed in L2 and L3 stage larvae committed to reproductive development and it is suppressed in dauers (DEpiFIG 5) (Gerisch et al., 2001; Mak and Ruvkun, 2004). Expresion of daf-9 solely in the hypodermis is sufficient to prevent dauer arrest and promote reproductive development (Gerisch and Antebi 2004; Mak and Ruvkun, 2004).
DEpiFIG 5: Hormonal regulation of dauer arrest by hypodermal DAF-9 activity. DAF-9 is an enzyme within in the biosynthentic pathway for dafachronic acid, the DAF-12/NHR ligand, and a major site for its function is in the hypodermis. A. Illustration shows expression of daf-9 in the hypodermis in larvae committed to reproductive development but not in dauers. Grey-filled drawings indicate that daf-9 is expressed, while white-filled ones indicate daf-9 is not expressed. B. When liganded, DAF-12 promotes reproductive development in target cells such as the somatic gonad. In the absence of ligand, DAF-12 complexed to the DIN-1 corepressor promotes dauer formation. |
4 References
Antebi, A., Culotti, J.G. and E.M. Hedgecock. 1998. daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development. 125: 1191-1205. Article
Antebi, A., Yeh, W.H., Tait, D., Hedgecock, E.M. and Riddle, D.L. 2000. daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 14: 1512-1527. Article
Burnell, A.M., Houthoofd, K., O’Hanlon, K. and Vanfleteren, J.R. 2005. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp. Geront. 40: 850-856. Abstract
Gerisch, B. and Antebi, A. 2004. Hormonal signals produced by DAF-9 cytochrome P450 regulate C. elegans dauer diapause in response to environmental cues. Development 131: 1765-1776. Article
Gerisch, B., Weitzel, C., Kober-Eisermann, C., Rottiers, V. and Antebi, A. 2001. A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell 1: 841-851. Article
Held, J.M., White, M.P., Fisher, A.L., Gibson, B.W., Lithgow, G.J. and Gill, M.S. 2006. DAF-12-dependent rescue of dauer formation in Caenorhabditis elegans by (25S)-cholestenoic acid. Aging Cell 5: 283-291. Article
Jia, K., Albert, P.S. and Riddle, D.L. 2002. DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 129: 221-23. Article
Liu, Z.C. and Ambros, V. 1989. Heterochronic genes control the stage-specific initiation and expression of the dauer larva developmental program in Caenorhabditis elegans. Genes Dev. 3: 2039-2049. Article
Ludewig, A.H., Kober-Eisermann, C., Weitzel, C., Bethke, A., Neubert, K., Gerisch, B., Hutter, H. and Antebi, A. 2004. A novel nuclear receptor/coregulator complex controls C. elegans lipid metabolism, larval development, and aging. Genes Dev. 18: 2120-2133. Article
Mak, H.Y. and Ruvkun, G. 2004. Intercellular signaling of reproductive development by the C. elegans DAF-9 cytochrome P450. Development 131: 1777-1786. Article
Meléndez, A., Tallóczy, Z., Seaman, M., Eskelinen, E L., Hall, D H., and Levine, B. 2003. Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science 301: 1387-1391. Abstract
Motola, D.L., Cummins, C.L., Rottiers, V., Sharma, K.K., Li, T., Li, Y., Suino-Powell, K., Xu, H.E., Auchus, R.J., Antebi, A. and Mangelsdorf, D.J. 2006. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 124:1209-1223. Article
Popham, J.D. and Webster, J.M. 1979. Aspects of the fine structure of the dauer larva of the nematode Caenorhabditis elegans. Can. J. Zool. 57: 794-800. Abstract
Schaedel, O.N., Gerisch, B., Antebi, A. and Sternberg, P.W. 2012. Hormonal signal amplification mediates environmental conditions during development and controls an irreversible commitment to adulthood. PLoS Biol. 10: e1001306. Article
Singh, R.N. and Sulston, J.E. 1978. Some observations on molting in Caenorhabditis elegans. Nematologica. 24: 63-71. Abstract
Vella, M.C. and Slack, F.J. 2005. C. elegans microRNAs (September 21, 2005). WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.26.1, http://www.wormbook.org. Article
Vowels, J.J. and Thomas, J.H. 1992. Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 130:105-123. Article
|

This chapter should be cited as: Wolkow, C.A. and Hall, D.H. 2015. The Dauer Epithelial System. In WormAtlas. doi:10.3908/wormatlas.3.9
Edited for the web by Laura A. Herndon. Last revision: March 26, 2015. |

|
|
|
|