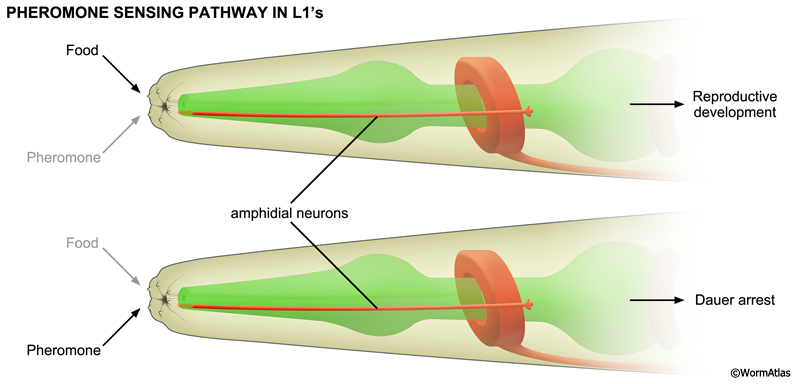
3 Neuronal Pathways Regulating Dauer Arrest
The dauer-inducing ascarosides act on amphid neurons that contact the environment at the distal openings of the amphid channels. Mutations causing amphid channel blockade impair pheromone signaling and prevent dauer arrest in response to pheromone (Albert et al., 1981). daf-7 encodes a TGF-beta ligand expressed in amphids that is critical for promoting reproductive development. daf-7 expression in ASI prevents dauer arrest and promotes development into fertile adults. Pheromone repressed the expression of daf-7::GFP reporters in ASI (Ren et al., 1996; Schackwitz et al., 1996; Peckol et al., 2001; Nolan, et al., 2002). Furthermore, ablation of ASI, together with ADF, ASG and ASJ, causes inappropriate dauer arrest under replete conditions (Bargmann and Horvitz, 1991; Schackwitz et al., 1996). Thus, pheromone directs dauer formation by repressing daf-7 pathway signaling in amphids.
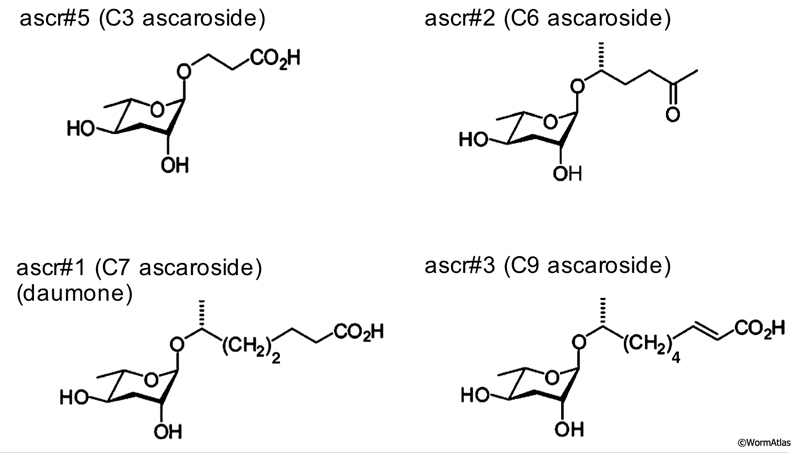
In the amphids, ascarosides signal via 7-transmembrane domain receptors coupled to G-protein signaling pathways. The receptors and signaling partners for C. elegans ascarosides appear to be ascaroside- and neuron-specific. In particular, ascr#2 (C6) and ascr#5 (C3) act on ASI through different receptors. In ASI, ascr#2 (C6) binds to DAF-37/GPCR, possibly in a heterodimeric complex with DAF-38, to directly repress daf-7 expression and promote dauer formation (Park et al., 2012). In contrast, ascr#5 (C3) binds to SRG-36/-37 heterodimeric receptors in ASI (McGrath et al., 2011).
Amphid signaling can affect dauer formation in other ways, as well. ASK activity seems to be required for larvae to enter the dauer stage, as ASK ablation can prevent dauer entry in the presence of pheromone (Schackwitz et al., 1996; Kim et al., 2009). In ASK neurons, ascr#2 (C6) signaling is mediated by the SRBC-64/-66 receptor, coupled to GPA-3/G-alpha, to inhibit DAF-11/guanylyl cyclase (Kim et al., 2009). It is thought that ASK controls dauer formation indirectly by acting on ASI through as yet undefined pathways (Kim et al., 2009). Finally, ASJ has been shown by ablation and exogenous activation experiments to promote dauer recovery (Bargmann and Horvitz, 1991; Schultheis et al., 2011). Since the food signal also promotes dauer recovery, ASJ may represent the site of action for this pathway (Golden and Riddle, 1984).
An insulin-like signaling pathway collaborates with the daf-7/TGF-beta pathway to regulate dauer formation (Morris et al., 1996; Kimura et al., 1997). The C. elegans genome encodes 40 insulin-like peptides (ILP), which are potential ligands for the DAF-2/insulin-receptor like protein (Pierce et al., 2001). ILPs are expressed throughout the body, including in neurons and the intestine. One neuronal ILP, DAF-28, expressed in ASI and ASJ, can antagonize dauer formation, suggesting it may transduce environmental dauer-inducing signals (Li et al., 2003). However, other ILPs impact daf-2 pathway activity from non-neuronal tissues, such as the intestine (Murphy et al., 2007).
Specialized endocrine cells in the head, named XXX cells (see Atypical Epithelial Cells), also regulate the dauer decision through synthesis of dafachronic acid (DA), the ligand for the DAF-12/nuclear hormone receptor (Gerisch et al., 2001; Jia et al., 2002). In L1 larvae, DA synthesis by the XXX cells reflects the presence or absence of environmental cues for dauer arrest, such as pheromone, food and temperature. In L1 larvae, favorable environmental conditions stimulate DA synthesis by the XXX cells, allowing larvae to bypass dauer. In later larval stages, the hypodermis becomes a second site for DA synthesis, propelling development into fertile adulthood (Schaedel et al., 2012). In unfavorable environments, DA synthesis by the XXX cells is reduced, committing larvae to enter dauer arrest (Schaedel et al., 2012). Components of the DAF-2/insulin-like pathway are also expressed in XXX cells, where they may modulate DA synthesis in response to environmental factors (Ohkura et al., 2003; Hu et al, 2006).
4 References
Ailion, M., Inoue, T., Weaver, C.I., Holdcraft, R.W. and Thomas, J.H. 1999. Neurosecretory control of aging in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 96: 7394-7397. Article
Ailion, M. and Thomas, J.H. 2003. Isolation and characterization of high-temperature-induced Dauer formation mutants in Caenorhabditis elegans. Genetics 165:127-144. Article
Albert, P.S., Brown, S.J. and Riddle, D.L. 1981. Sensory control of dauer larva formation in Caenorhabditis elegans. J. Comp. Neurol. 198: 435-451. Abstract
Bargmann, C.I. and Horvitz, H.R. 1991. Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science 251: 1243-1246. Abstract
Braendle, C. 2012. Pheromones: Evolving language of chemical communication in nematodes. Curr. Biol. 22: R294-R296. Abstract
Butcher, R.A., Fujita, M., Schroeder, F.C. and Clardy, J. 2007. Small-molecule pheromones that control development in Caenorhabditis elegans. Nat. Chem. Biol. 3: 420-422. Abstract
Butcher, R.A., Ragains, J.R., Kim, E. and Clardy, J. 2008. A potent dauer pheromone component in Caenorhabditis elegans that acts synergistically with other components. Proc. Natl. Acad. Sci. 105: 14288-14292. Article
Butcher, R.A., Ragains, J.R., Li, W., Ruvkun, G., Clardy, J. and Mac, H.Y. 2009. Biosynthesis of the Caenorhabditis elegans dauer pheromone. Proc. Natl. Acad. Sci. 106: 1875-1879. Article
Choe, A., von Reuss, S.H., Kogan, D., Gasser, R.B., Platzer, E.G., Schroeder, F.C. and Sternberg, P. 2012a. Ascaroside signaling is widely conserved among nematodes. Curr. Biol. 22: 772-780. Abstract
Choe, A., Chuman, T., von Reuss, S.H., Dossey, A.T., Yim, J.J., Ajredini, R., Kolawa, A.A., Kaplan, F., Alborn, H.T., Teal, P.E.A., Schroeder, F.C., Sternberg, P.W. and Edison, A.S. 2012b. Sex-specific mating pheromones in the nematode Panagrellus redivivus. Proc. Natl. Acad. Sci. 109: 20949-20954. Abstract
Gerisch, B., Weitzel, C., Kober-Eisermann, C., Rottiers, V. and Antebi, A. 2001. A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell. 1: 841-851. Article
Golden, J.W. and Riddle, D.L. 1982. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science. 218: 578-580. Abstract
Golden, J.W. and Riddle, D.L. 1984. Caenorhabditis elegans dauer larva: Developmental effects of pheromone, food and temperature. Dev. Biol. 102: 368-378. Abstract
Hu, P.J., Xu, J. and Ruvkun, G. 2006. Two membrane-associated tyrosine phosphatase homologs potentiate C. elegans AKT-1/PKB signaling. PLoS Genet. 2: e99. Article
Jeong, P.-Y., Jung, M., Yim, Y.-H., Kim, H., Park, M., Hong, E., Lee, W., Kim, Y.H., Kim, K. and Paik, Y.-K. 2005. Chemical structure and biological activity of the Caenorhabditis elegans dauer-inducing pheromone. Nature 433: 541-545. Abstract
Jia, K., Albert, P.S. and Riddle, D.L. 2002. DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 129: 221-231. Article
Joo, H.-J., Yim, Y.-H., Jeong, P.-Y., Jin, Y.-X., Lee, J.-E., Kim, H., Jeong, S.-K., Chitwood, D.J. and Paik, Y.-K. 2009. Caenorhabditis elegans utilizes dauer pheromone biosynthesis to dispose of toxic peroxisomal fatty acids for cellular homeostasis. Biochem. J. 422: 61-71. Article
Joo, H.-J., Kim, K.-Y., Yim, Y.-H., Jin, Y.-X., Kim, H., Kim, M.-Y. and Paik, Y.-K. 2010. Contribution of the peroxisomal acox gene to the dynamic balance of daumone production in Caenorhabditis elegans. J. Biol. Chem. 285: 29319-29325. Article
Kim, K., Sato, K., Shibuya, M., Zeiger, D.M., Butcher, R.A., Ragains, J.R., Clardy, J., Touhara, K., and Sengupta, P. 2009. Two chemoreceptors mediate developmental effects of dauer pheromone in C. elegans. Science 326: 994-998. Abstract
Kimura, K.D., Tissenbaum, H.A., Liu, Y., and Ruvkun, G. 1997. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277: 942-946. Abstract
Li, W., Kennedy, S.G. and Ruvkun, G. 2003. daf-28 encodes a C. elegans insulin superfamily member that is regulated by environmental cues and acts in the DAF-2 signaling pathway. Genes Dev. 17: 844-858. Article
Ludewig, A.H. and Schroeder, F.C. 2013. Ascaroside signaling in C. elegans. In WormBook (ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.155.1) Article
Mahoney, T.R., Liu, Q., Itoh, T., Luo, S., Hadwiger, G., Vincent, R., Wang, Z-W., Fukuda, M. and Nonet, M.L. 2006. Regulation of synaptic transmission by RAB-3 and RAB-27 in Caenorhabditis elegans. Mol. Biol. Cell 17: 2617-2625. Article
McGrath, P.T., Xu, Y., Ailion, M., Garrison, J.L., Butcher, R.A. and Bargmann, C.I. 2011. Parallel evolution of domesticated Caenorhabditis species targets pheromone receptor genes. Nature 477: 321-325. Abstract
Morris, J.Z., Tissenbaum, H.A., and Ruvkun, G. 1997. A phosphatidylinositol-3-OH kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 382: 536-539. Abstract
Murphy, C.T., Lee, S.-J. and Kenyon, C. 2007. Tissue entrainment by feedback regulation of insulin gene expression in the endoderm of Caenorhabditis elegans. Proc. Natl. Acad. Sci. 104: 19046-50. Article
Noguez, J.H., Conner, E.S., Zhou, Y., Ciche, T.A., Ragains, J.R. and Butcher, R.A. 2012. A novel ascaroside controls the parasitic life cycle of the entomopathogenic nematode Heterorhabditis bacteriophora. ACS Chem. Biol. 7: 961-966. Abstract
Nolan, K.M., Sarafi-Reinach, T.R., Horne, J.G., Saffer, A.M. and Sengupta, P. 2002. The DAF-7 TGF-beta signaling pathway regulates chemosensory receptor gene expression in C. elegans. Genes Dev. 16: 3061-73. Article
Ohkura, K., Suzuki, N., Ishihara, T. and Katsura, I. 2003. SDF-9, a protein tyrosine phosphatase-like molecule, regulates the L3/dauer developmental decision through hormonal signaling in C. elegans. Development 14: 3237-3248. Article
Park, D., O’Doherty, I., Somvanshi, R.K., Bethke, A., Schroeder, F.C., Kumar, U. and Riddle, D.L. 2012. Interaction of structure-specific and promiscuous G-protein-coupled receptors mediates small-molecule signaling in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 109: 9917-9922. Abstract
Peckol, E.L, Troemel, E.R. and Bargmann, C.I. 2001. Sensory experience and sensory activity regulate chemosensory receptor gene expression in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 98: 11032-11038. Article
Pierce, S.B., Costa, M., WIsotzkey, R., Devadhar, S., Homburger, S.A., Buchman, A.R., Ferguson, K.C., Heller, J., Platt, D.M., Pasquinelli, A.A., Liu, L.X., Doberstein, S.K. and Ruvkun, G. 2001. Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev. 15: 672-86. Article
Ren, P., Lim, C.S., Johnsen, R., Albert, P.S., Pilgrim, D, and Riddle, D.L. 1996. Control of C. elegans larval development by neuronal expression of a TGF-β homolog. Science 274: 1389-1391. Abstract
Schackwitz, W.S., Inoue, T. and Thomas, J.H. 1996. Chemosensory neurons function in parallel to mediate a pheromone response in C. elegans. Neuron 17: 719-728. Article
Schaedel, O.N., Gerisch, B., Antebi, A. and Sternberg, P.W. 2012. Hormonal signal amplification mediates environmental conditions during development and controls an irreversible commitment to adulthood. PLoS Biol. 10: e1001306. Article
Schultheis, C., Liewald, J.F., Bamberg, E., Nagel, G. and Gottschalk, A. 2011. Optogenetic long-term manipulation of behavior and animal development. PLoS ONE. 6: e18766. Article
von Reuss, S.H., Bose, N., Srinivasan, J., Yim, J.J., Judkins, J.C., Sternberg, P.W. and Schroeder, F.C. 2012. Comparative metabolomics reveals biogenesis of ascarosides, a modular library of small-molecule signals in C. elegans. J. Am. Chem. Soc. 134: 1817-1824. Article
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image