|
The Pharynx of
Caenorhabditis elegans
Donna G. Albertson and J.N. Thomson
M.R.C. Laboratory of Molecular Biology
Hills Road, Cambridge, CB2 2QH, England
Phil. Trans. Royal Soc. London (1976) 275B: 299-325
doi: 10.1098/rstb.1976.0085
(Communicated by S. Brenner, F.R.S. Received 15 September 1975)
Abstract - Introduction -
Material & Methods -
Results -
Discussion -
Acknowledgments
- References
Abstract
The anatomy of the pharynx of Caenorhabditis elegans has been reconstructed from electron micrographs of serial sections. The pharynx is used for pumping food into the gut, and is composed of 34 muscle cells, 9 marginal cells, 9 epithelial cells, 5 gland cells and 20 neurones. Three regions of specialization in the cuticle lining of the pharyngeal lumen may aid in the accumulation of food particles. A basement membrane isolates the pharynx from the rest of the animal, making the pharyngeal nervous system a nearly self-contained unit which is composed primarily of five classes of motor neurones and six classes of interneurones. Three other classes have also been described, which by their morphology appear to be neurosecretory and motor, motor and interneuronal, and lastly one pair that only innervates three of the marginal cells. Some classes of neurone have free endings just under the cuticle lining the lumen of the pharynx, suggesting that these are mechano- or proprio-receptive endings. The connectivity of these neurones has been described at the level of individual synaptic regions, and after combining this information with video taped observations of the pharynx pumping, some interpretations of how these neurones function have been offered.
Introduction
The nematode Caenorhabditis elegans is a favourable organism for the study of simple nervous systems because it is amenable to ultrastructural and genetic analyses (Brenner 1973; Ward, Thomson, White & Brenner 1975; Ware, Clark, Crossland & Russell 1975; White, Southgate, Thomson & Brenner 1976 ; Sulston 1976). In this paper we report the detailed structure of the pharynx.
The pharynx is a muscular pump and is a prominent feature of the nematode's head. Rhythmic contractions of the seven muscle regions suck bacteria into the lumen, grind them and pass them back into the gut. In addition, the structure contains epithelial, and gland cells. Its own nervous system, composed of twenty cells, is nearly completely isolated from the somatic nervous system.
Thus the pharynx is a self-contained neuromuscular system that is small enough to be described in detail by reconstruction from electron micrographs of serial sections. When combined with behavioural observations, this information might be used to make some suggestions about the functioning of simple nervous systems.
Materials & Methods
Preparation and electron microscopy of 50 nm thick serial sections through Caenorhabditis elegans have been described (Ward et al. 1975; White et al. 1976, following paper).
The anatomy of the pharynx was reconstructed by tracing cell sections on electron micrographs in the same manner as described by Ward et al. (1975). A single series of one animal included the entire pharynx. Most cells in the pharynx are large and easily reconstructed from photographs taken of every third section, but in the region of the nerve ring in the metacorpus and where a commissure is found in the terminal bulb, additional series were necessary. At each of these places a series collected on slot grids was prepared so that all sections were visible for tracing neurone processes. Except in the middle of the procorpus, at least one additional partial series was cut for conformation of the findings. The distinctive morphology of neurones in the pharynx allows one to identify cell processes when their cell bodies are not included within a partial series. Further checks on assignments are made by comparison of bilaterally symmetric pairing of cells or cell processes.
Computer aided reconstruction from electron micrographs used in the preparation of some drawings has been described (White et al. 1976).
Results
(a) General plan of the pharynx
(b) Cuticle
(c) Marginal cells
(d) Gland cells
(e) Pharyngeal-intestinal valve
(f) Muscles
(g) Nervous system
(i) Topography of the nervous system
(ii) Connectivity
A. Input to the pharyngeal nervous system
B. Pharyngeal connectivity
(iii) Variability
(a) General plan of the pharynx
The pharynx of C. elegans is two-lobed, or of the rhabditoid type (Chitwood & Chitwood 1938). Photographed in the living animal in figure 1, plate 1, one can easily distinguish the buccal cavity, procorpus, metacorpus, isthmus, and terminal bulb. A basement membrane completely surrounds the pharynx and isolates it from the rest of the animal.
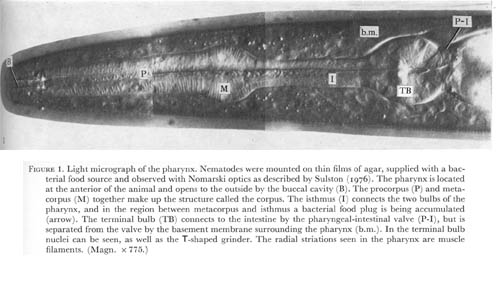 Figure 1. Plate 1 - Light micrograph of the pharynx. Nematodes were mounted on thin films of agar, supplied with a bacterial food source and observed with Nomarski optics as described by Sulston (1976). The pharynx is located at the anterior of the animal and opens to the outside by the buccal cavity (B). The procorpus (P) and metacorpus (M) together make up the structure called the corpus. The isthmus (I) connects the two bulbs of the pharynx, and in the region between metacorpus and isthmus a bacterial food plug is being accumulated (arrow). The terminal bulb (TB) connects to the intestine by the pharyngeal-intestinal valve (P-I), but is separated from the valve by the basement membrane surrounding the pharynx (b.m.). In the terminal bulb nuclei can be seen, as well as the T-shaped grinder. The radial striations seen in the pharynx are muscle filaments. (Magn. x 775.)
Figure 1. Plate 1 - Light micrograph of the pharynx. Nematodes were mounted on thin films of agar, supplied with a bacterial food source and observed with Nomarski optics as described by Sulston (1976). The pharynx is located at the anterior of the animal and opens to the outside by the buccal cavity (B). The procorpus (P) and metacorpus (M) together make up the structure called the corpus. The isthmus (I) connects the two bulbs of the pharynx, and in the region between metacorpus and isthmus a bacterial food plug is being accumulated (arrow). The terminal bulb (TB) connects to the intestine by the pharyngeal-intestinal valve (P-I), but is separated from the valve by the basement membrane surrounding the pharynx (b.m.). In the terminal bulb nuclei can be seen, as well as the T-shaped grinder. The radial striations seen in the pharynx are muscle filaments. (Magn. x 775.)
The lumen of the pharynx and buccal cavity are lined with cuticle. Anteriorly, the lumen is triangular, but in the procorpus it becomes triradiate, with tubular apices. Further posterior in the procorpus it is completely closed. The metacorpus is slightly more open and the tubular apices reappear briefly at the anterior of the isthmus. Three specialized regions are found, one at the beginning of the procorpus, one at the back of the metacorpus and one in the terminal bulb. These will be described separately.
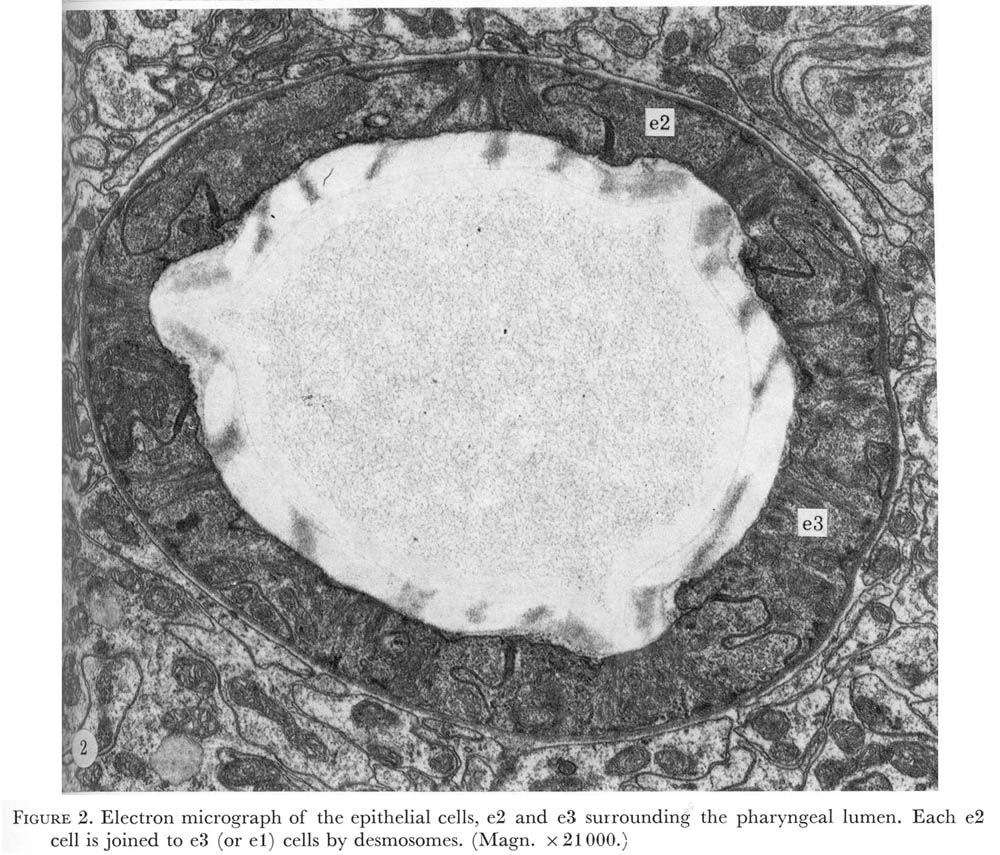
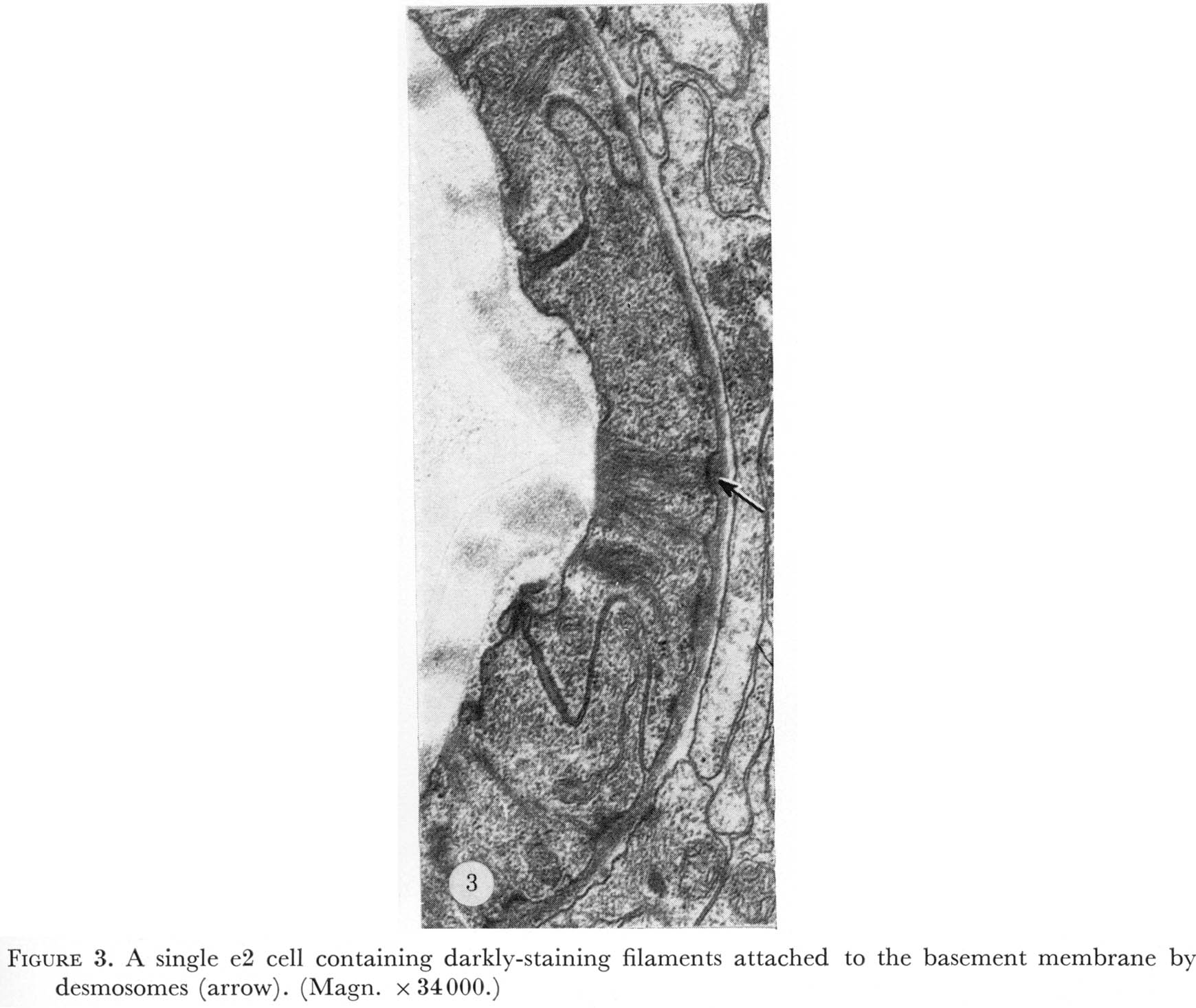
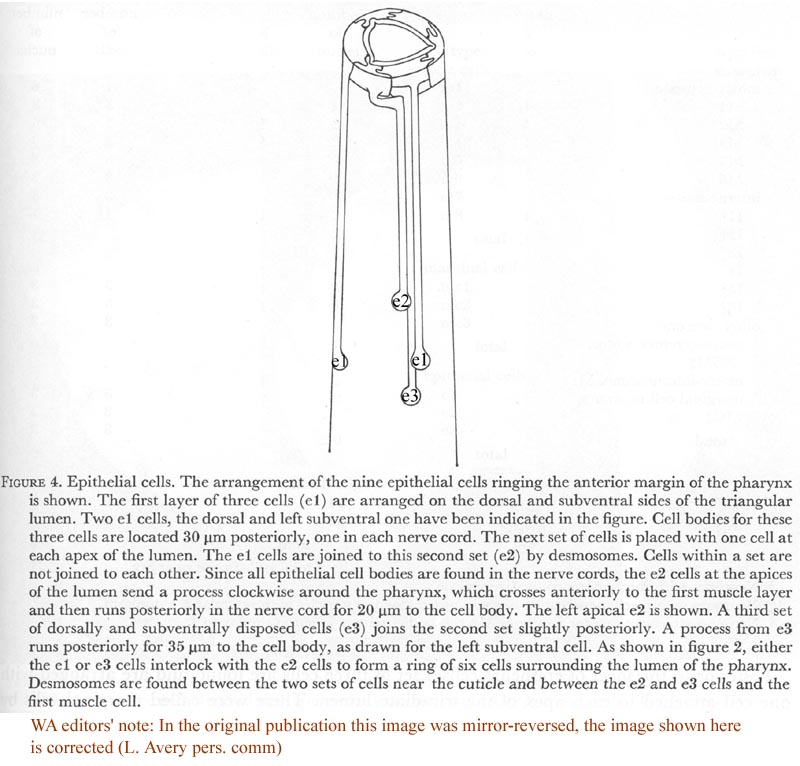
Nine epithelial cells ring the anterior margin of the pharynx (figure 2, plate 1). The cells contain filaments that extend from the pharyngeal cuticle to the basement membrane. The attachment of the filaments to the basement membrane have been called half-desmosomes (Lee 1968) and are characterized by enlargement and darkening of the interior margin of the basement membrane as shown in cross section in figure 3, plate 1. As shown in figure 4 these divide into three sets of three cells each. The anterior six cells project slightly into the arcade cells, which lie along the outside of the pharynx in the nematode's head (Ward et al. 1975), these cells stitch the pharynx to the arcade cells by desmosomes.
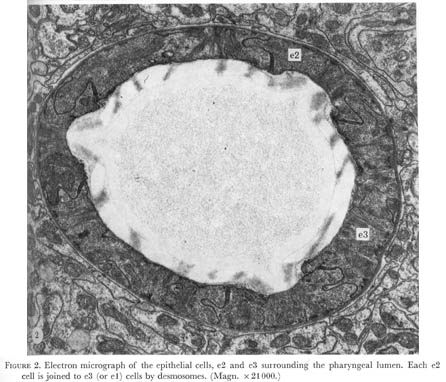 Figure 2. Plate 1 - Electron micrograph of the epithelial cells, e2 and e3 surrounding the pharyngeal lumen. Each e2 cell is joined to e3 (or e1) cells by desmosomes. (Magn. x 21000.)
Figure 2. Plate 1 - Electron micrograph of the epithelial cells, e2 and e3 surrounding the pharyngeal lumen. Each e2 cell is joined to e3 (or e1) cells by desmosomes. (Magn. x 21000.)
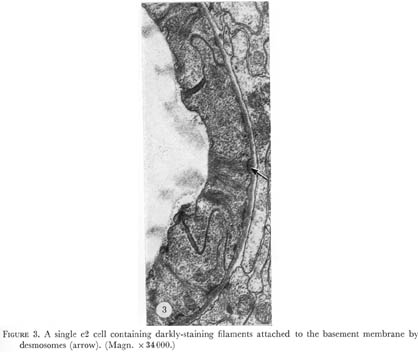 Figure 3. Plate 1 - A single e2 cell containing darkly-staining filaments attached to the basement membrane by half desmosomes (arrow). (Magn. x 34000.)
Figure 3. Plate 1 - A single e2 cell containing darkly-staining filaments attached to the basement membrane by half desmosomes (arrow). (Magn. x 34000.)
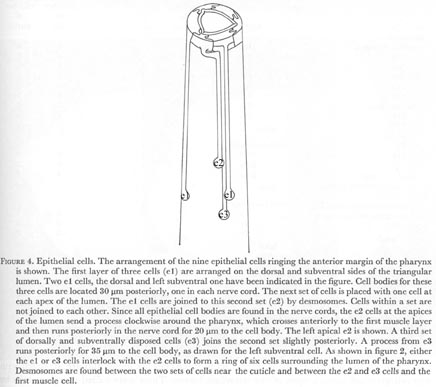 Figure 4. Epithelial cells. The arrangement of the nine epithelial cells ringing the anterior margin of the pharynx is shown. The first layer of three cells (e1) are arranged on the dorsal and subventral sides of the triangular lumen. Two e1 cells, the dorsal and left subventral one have been indicated in the figure. Cell bodies for these three cells are located 30 um (micro) posteriorly, one in each nerve cord. The next set of cells is placed with one cell at each apex of the lumen. The e1 cells are joined to this second set (e2) by desmosomes. Cells within a set are not joined to each other. Since all epithelial cell bodies are found in the nerve cords, the e2 cells at the apices of the lumen send a process clockwise around the pharynx, which crosses anteriorly to the first muscle layer and then runs posteriorly in the nerve cord for 20 um to the cell body. The left apical e2 is shown. A third set of dorsally and subventrally disposed cells (e3) joins the second set slightly posteriorly. A process from e3 runs posteriorly for 35 um to the cell body, as drawn for the left subventral cell. As shown in figure 2, either the e1 or e3 cells interlock with the e2 cells to form a ring of six cells surrounding the lumen of the pharynx. Desmosomes are found between the two sets of cells near the cuticle and between the e2 and e3 cells and the first muscle cell.
Figure 4. Epithelial cells. The arrangement of the nine epithelial cells ringing the anterior margin of the pharynx is shown. The first layer of three cells (e1) are arranged on the dorsal and subventral sides of the triangular lumen. Two e1 cells, the dorsal and left subventral one have been indicated in the figure. Cell bodies for these three cells are located 30 um (micro) posteriorly, one in each nerve cord. The next set of cells is placed with one cell at each apex of the lumen. The e1 cells are joined to this second set (e2) by desmosomes. Cells within a set are not joined to each other. Since all epithelial cell bodies are found in the nerve cords, the e2 cells at the apices of the lumen send a process clockwise around the pharynx, which crosses anteriorly to the first muscle layer and then runs posteriorly in the nerve cord for 20 um to the cell body. The left apical e2 is shown. A third set of dorsally and subventrally disposed cells (e3) joins the second set slightly posteriorly. A process from e3 runs posteriorly for 35 um to the cell body, as drawn for the left subventral cell. As shown in figure 2, either the e1 or e3 cells interlock with the e2 cells to form a ring of six cells surrounding the lumen of the pharynx. Desmosomes are found between the two sets of cells near the cuticle and between the e2 and e3 cells and the first muscle cell.
WA editors' note: In the original publication the location of the epithelial cells were mirror-reversed. The image shown here is the corrected version (L. Avery pers. comm.)
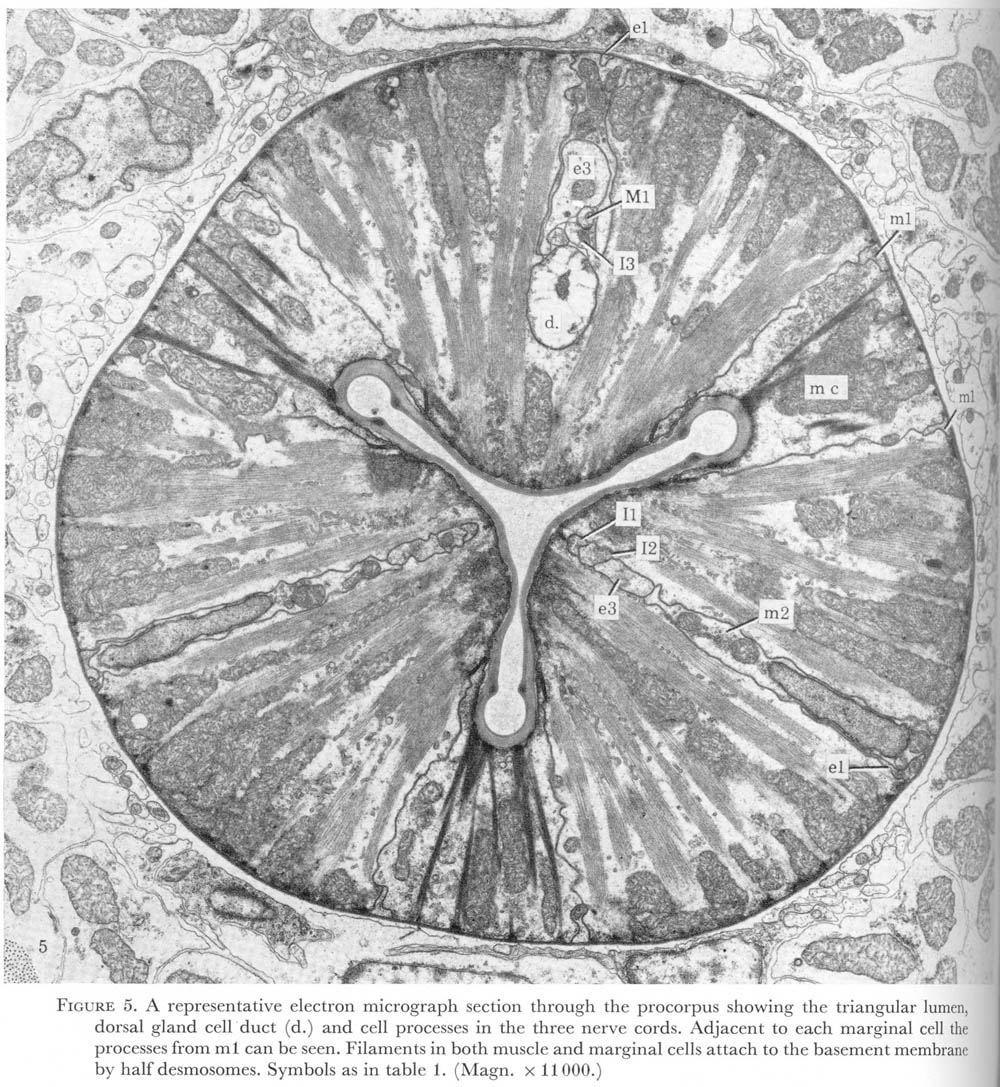
Posterior to the sheet of epithelial cells a set of three cells are found and are arranged with one cell attached to each apex of the triradiate lumen. These were called marginal cells by Looss (1905) as referenced in Bird 1971, and the dorsal and two subventral muscle sectors which they delineate from the general plan of the pharynx (figure 5, plate 2). In the middle of each muscle sector neuronal processes run, forming three cords, stretching from the anterior end of the pharynx to the terminal bulb. In the corpus the cords contain epithelial and some muscle cell nuclei, as well as the neurone cell bodies. Here also, the dorsal nerve cord is accompanied by the dorsal gland cell duct. In the isthmus the subventral glands lie next to the subventral cords, so that all three sectors contain ducts in this region (figure 6, plate 3).
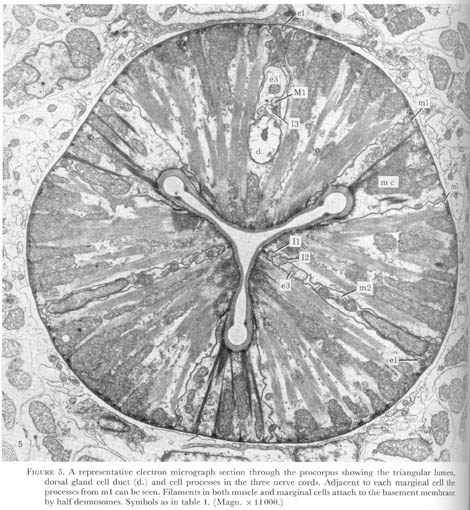 Figure 5. Plate 2 - A representative electron micrograph section through the pro corpus showing the triangular lumen, dorsal gland cell duct (d.) and cell processes in the three nerve cords. Adjacent to each marginal cell the processes from m1 can be seen. Filaments in both muscle and marginal cells attach to the basement membrane by halfdesmosomes. Symbols as in table 1. (Magn. x 11000.)
Figure 5. Plate 2 - A representative electron micrograph section through the pro corpus showing the triangular lumen, dorsal gland cell duct (d.) and cell processes in the three nerve cords. Adjacent to each marginal cell the processes from m1 can be seen. Filaments in both muscle and marginal cells attach to the basement membrane by halfdesmosomes. Symbols as in table 1. (Magn. x 11000.)
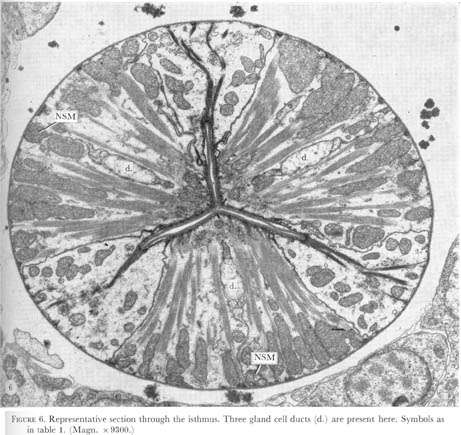 Figure 6. Plate 3 - Representative section through the isthmus.
Three gland cell ducts (d.) are present here. Symbols as in table 1. (Magn. x 9300.)
Figure 6. Plate 3 - Representative section through the isthmus.
Three gland cell ducts (d.) are present here. Symbols as in table 1. (Magn. x 9300.)
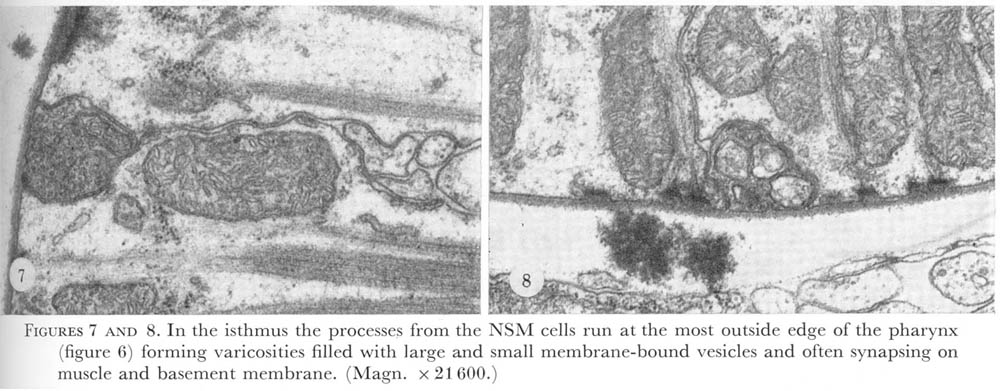
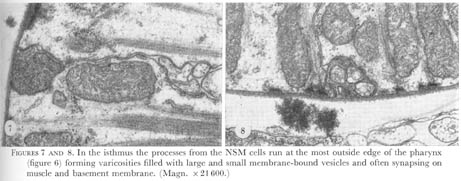 Figure 7&8. Plate 3 - In the isthmus the processes from the NSM cells run at the most outside edge of the pharynx (figure 6) forming varicosities filled with large and small membrane-bound vesicles and often synapsing on muscle and basement membrane. (Magn. X 21600.)
Figure 7&8. Plate 3 - In the isthmus the processes from the NSM cells run at the most outside edge of the pharynx (figure 6) forming varicosities filled with large and small membrane-bound vesicles and often synapsing on muscle and basement membrane. (Magn. X 21600.)
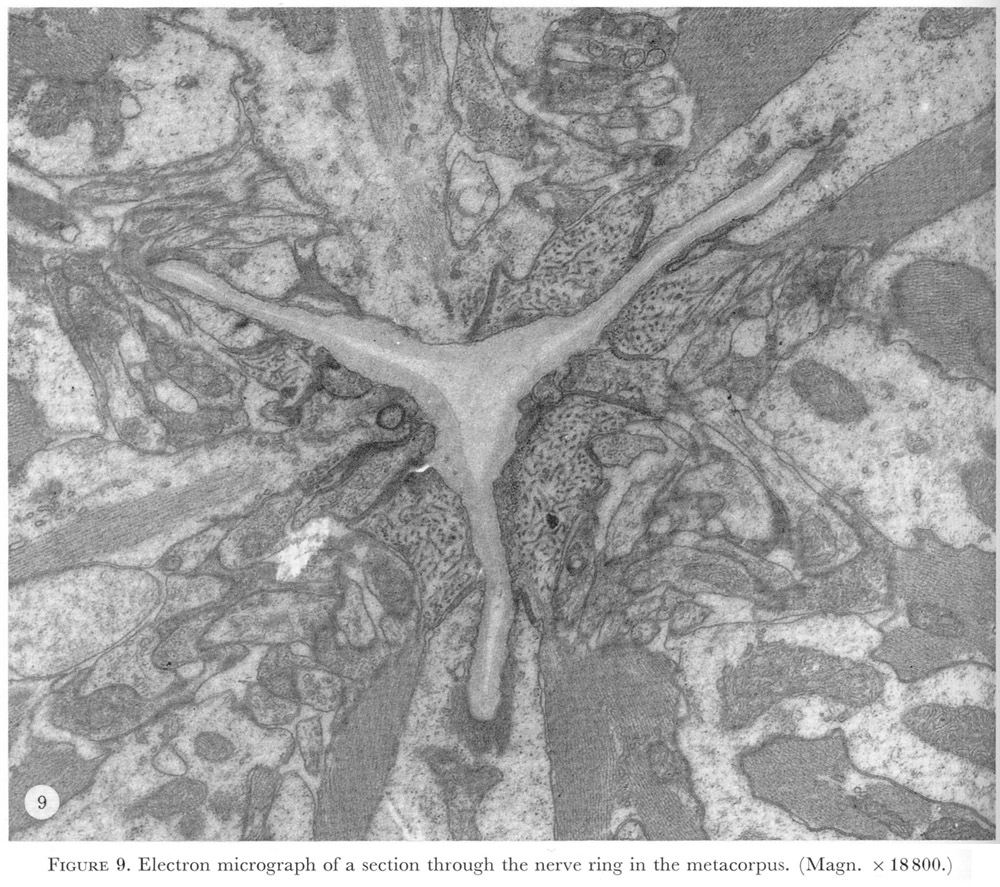
In the metacorpus nerve cell processes from the three cords in the procorpus and isthmus meet and form a complete nerve ring (figure 9, plate 4).
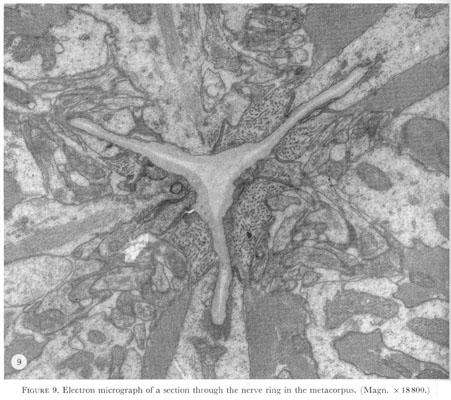 Figure 9. Plate 4 - Electron micrograph of a section through the nerve ring in the metacorpus. (Magn. x 18800.)
Figure 9. Plate 4 - Electron micrograph of a section through the nerve ring in the metacorpus. (Magn. x 18800.)
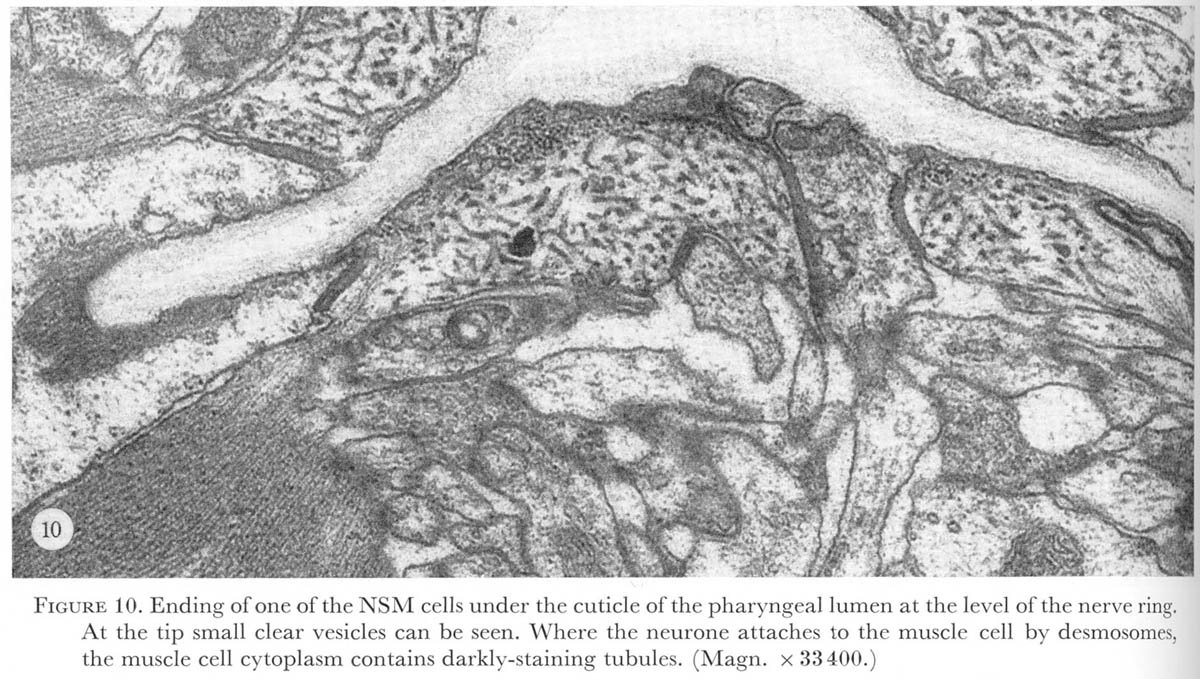
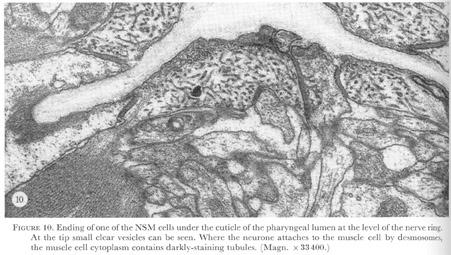 Figure 10. Plate 4 - Ending of one of the NSM cells under the cuticle of the pharyngeal lumen at the level of the nerve ring. At the tip small clear vesicles can be seen. Where the neurone attaches to the muscle cell by desmosomes, the muscle cell cytoplasm contains darkly-staining tubules. (Magn. X 33400.)
Figure 10. Plate 4 - Ending of one of the NSM cells under the cuticle of the pharyngeal lumen at the level of the nerve ring. At the tip small clear vesicles can be seen. Where the neurone attaches to the muscle cell by desmosomes, the muscle cell cytoplasm contains darkly-staining tubules. (Magn. X 33400.)
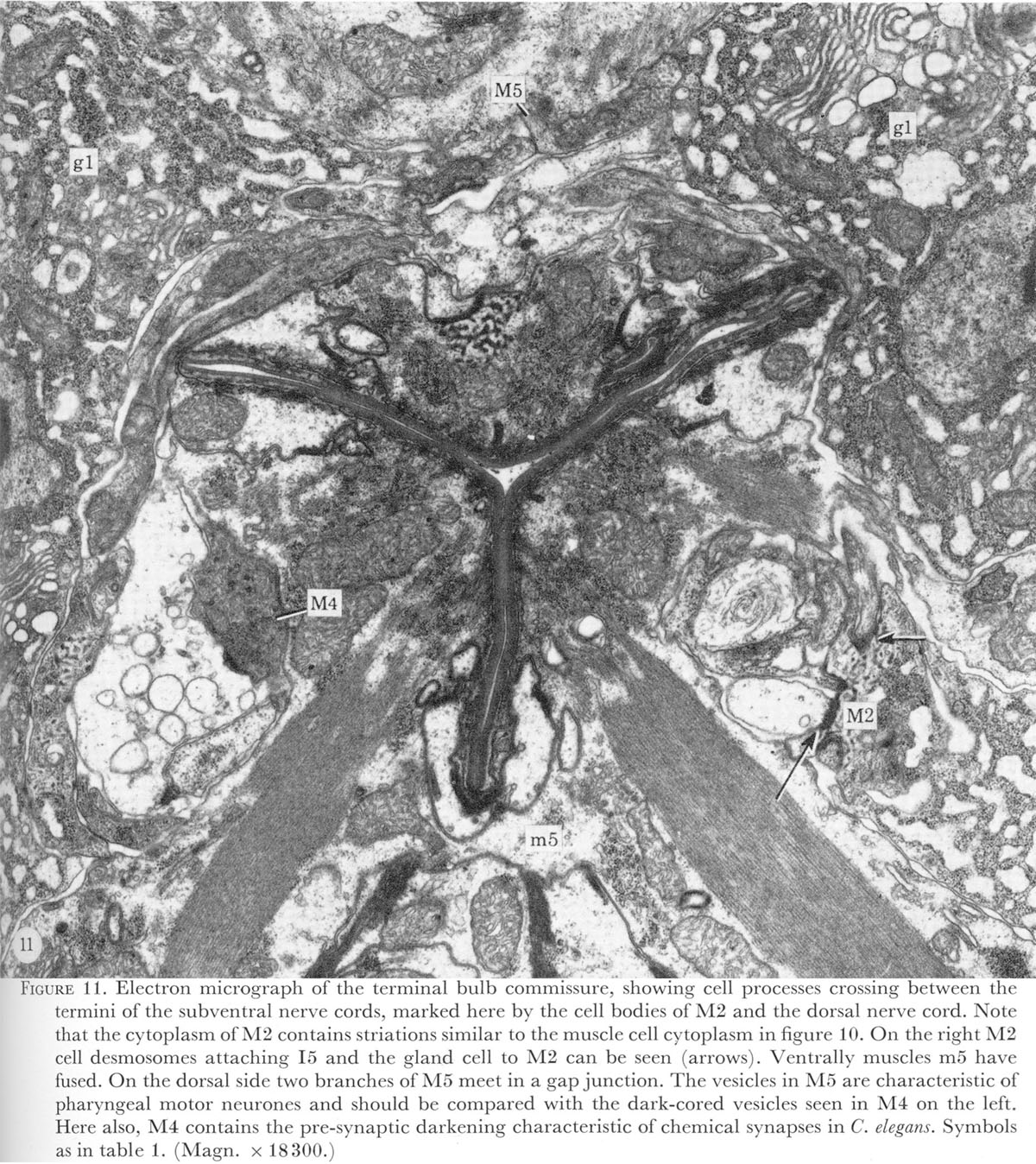
At the termination of the nerve cords a horseshoe-shaped terminal bulb commissure is formed by processes crossing between the subventral nerve cords in the isthmus and the dorsal cord. At this level too, the five gland cells become a prominent feature (figure 11, plate 5). The pharynx terminates in the pharyngeal-intestinal valve or cardia, which is seen in figure 1, plate 1, as a collar linking pharynx and intestine.
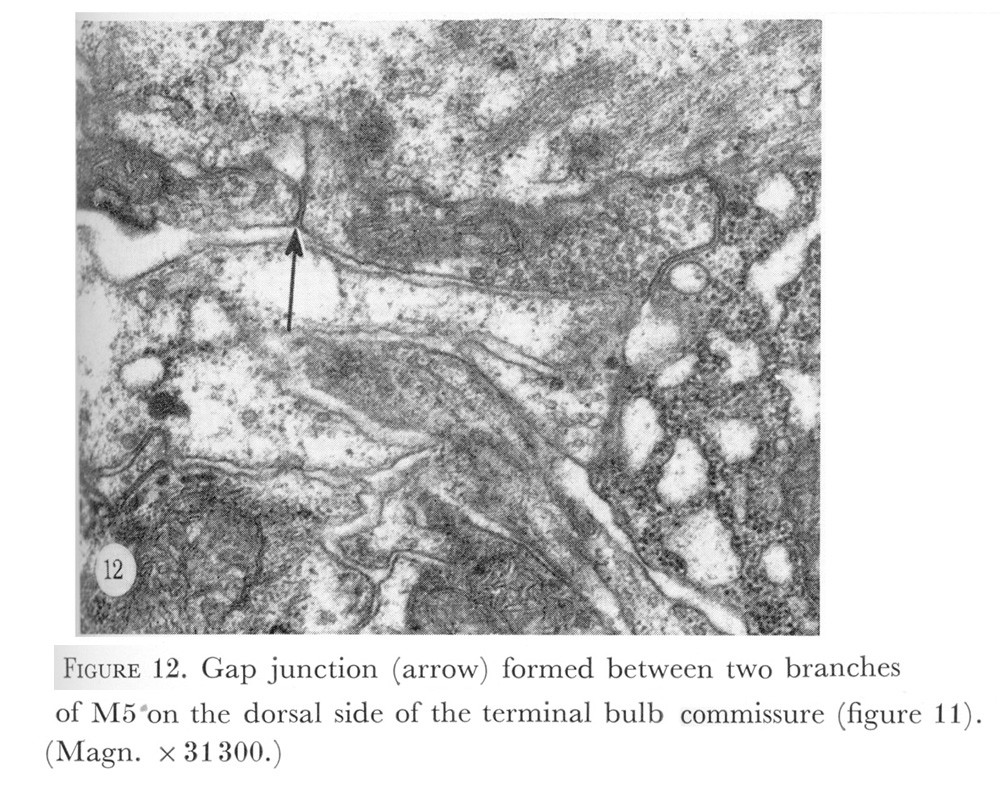
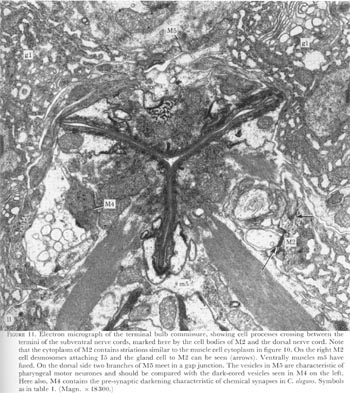 Figure 11. Plate 5 - Electron micrograph of the terminal bulb commissure, showing cell processes crossing between the termini of the subventral nerve cords, marked here by the cell bodies of M2 and the dorsal nerve cord. Note that the cytoplasm of M2 contains striations similar to the muscle cell cytoplasm in figure 10. On the right M2 cell desmosomes attaching I5 and the gland cell to M2 can be seen (arrows). Ventrally muscles m5 have fused. On the dorsal side two branches of M5 meet in a gap junction. The vesicles in M5 are characteristic of pharyngeal motor neurones and should be compared with the dark-cored vesicles seen in M4 on the left. Here also, M4 contains the pre-synaptic darkening characteristic of chemical synapses in C. elegans. Symbols as in table 1. (Magn. x 18300.)
Figure 11. Plate 5 - Electron micrograph of the terminal bulb commissure, showing cell processes crossing between the termini of the subventral nerve cords, marked here by the cell bodies of M2 and the dorsal nerve cord. Note that the cytoplasm of M2 contains striations similar to the muscle cell cytoplasm in figure 10. On the right M2 cell desmosomes attaching I5 and the gland cell to M2 can be seen (arrows). Ventrally muscles m5 have fused. On the dorsal side two branches of M5 meet in a gap junction. The vesicles in M5 are characteristic of pharyngeal motor neurones and should be compared with the dark-cored vesicles seen in M4 on the left. Here also, M4 contains the pre-synaptic darkening characteristic of chemical synapses in C. elegans. Symbols as in table 1. (Magn. x 18300.)
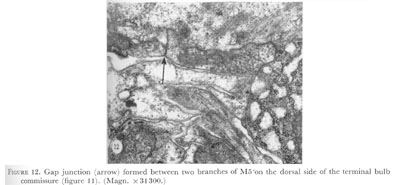 Figure 12. Plate 5 - Gap junction (arrow) formed between two branches of M5 on the dorsal side of the terminal bulb commissure (figure 11). (Magn. X 31300.)
Figure 12. Plate 5 - Gap junction (arrow) formed between two branches of M5 on the dorsal side of the terminal bulb commissure (figure 11). (Magn. X 31300.)
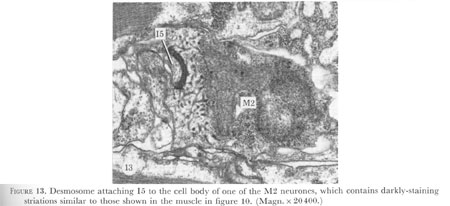 Figure 13. Plate 5 - Desmosome attaching I5 to the cell body of one of the M2 neurones, which contains darkly-staining striations similar to those shown in the muscle in figure 10. (Magn. X 20400.)
Figure 13. Plate 5 - Desmosome attaching I5 to the cell body of one of the M2 neurones, which contains darkly-staining striations similar to those shown in the muscle in figure 10. (Magn. X 20400.)
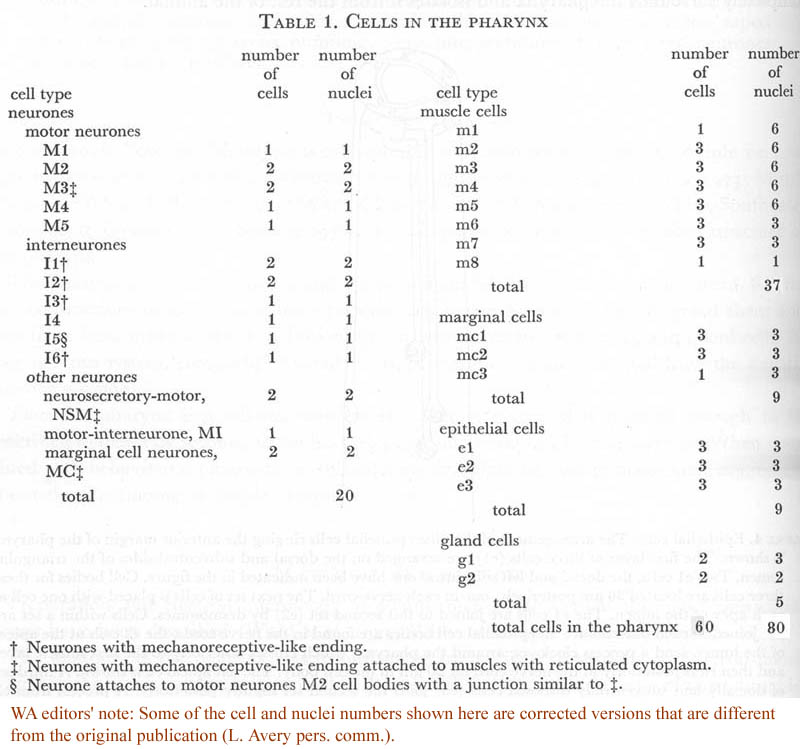
Further descriptions of pharyngeal structure will be presented in the following sections. Greatest attention has been given to the neural circuitry. Table 1 summarizes the number and classes of all pharyngeal cells, while figure 14 displays the positions of all nuclei in the pharynx.
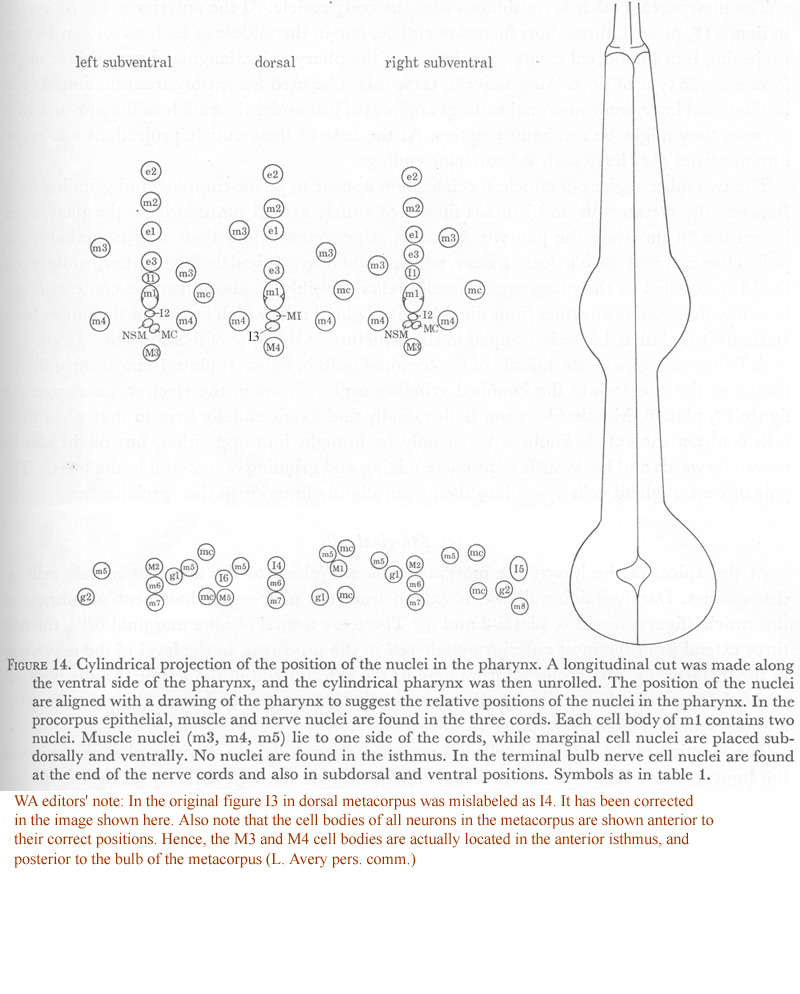
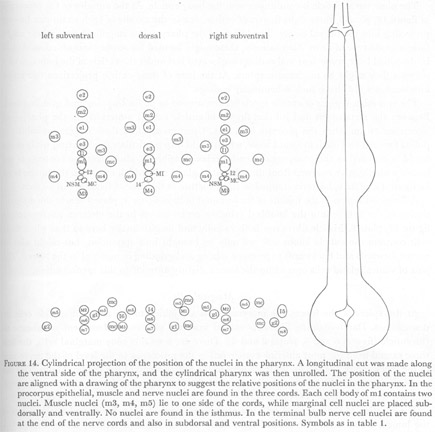 Figure 14. Cylindrical projection of the position of the nuclei in the pharynx. A longitudinal cut was made along the ventral side of the pharynx, and the cylindrical pharynx was then unrolled. The position of the nuclei are aligned with a drawing of the pharynx to suggest the relative positions of the nuclei in the pharynx. In the procorpus epithelial, muscle and nerve nuclei are found in the three cords. Each cell body of m1 contains two nuclei. Muscle nuclei (m3, m4, m5) lie to one side of the cords, while marginal cell nuclei are placed sub-dorsally and ventrally. No nuclei are found in the isthmus. In the terminal bulb nerve cell nuclei are found at the end of the nerve cords and also in subdorsal and ventral positions. Symbols as in table 1.
Figure 14. Cylindrical projection of the position of the nuclei in the pharynx. A longitudinal cut was made along the ventral side of the pharynx, and the cylindrical pharynx was then unrolled. The position of the nuclei are aligned with a drawing of the pharynx to suggest the relative positions of the nuclei in the pharynx. In the procorpus epithelial, muscle and nerve nuclei are found in the three cords. Each cell body of m1 contains two nuclei. Muscle nuclei (m3, m4, m5) lie to one side of the cords, while marginal cell nuclei are placed sub-dorsally and ventrally. No nuclei are found in the isthmus. In the terminal bulb nerve cell nuclei are found at the end of the nerve cords and also in subdorsal and ventral positions. Symbols as in table 1.
WA editors' note: In the original figure I3 neuron in dorsal metacorpus was mislabeled as I4. It has been corrected in the image shown here. Also note that the cell bodies of all neurons in the metacorpus are shown anterior to their correct positions in this image. Hence, the M3 and M4 cell bodies are actually located in the anterior isthmus and posterior to the bulb of the metacorpus.
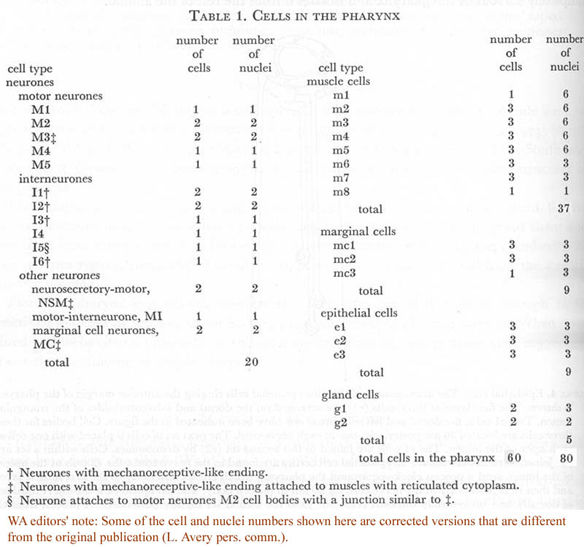 Table 1. Cells in the pharynx.
Table 1. Cells in the pharynx.
(b) Cuticle
The pharyngeal cuticle is continuous with the body cuticle. At the anterior of the procorpus in figure 15, plate 6, three short fingers of cuticle, one in the middle of each sector can be seen projecting into the buccal cavity. The lumen of the pharynx is triangular here so these might form a crude type of sieve. Alternatively, these might be used for sensory transduction. As will be described later, some neuronal endings are located just under the cuticle of the lumen, which suggests they might be mechanoreceptors. At the base of these cuticle projections two major interneurones (I2) have such subcuticular endings.
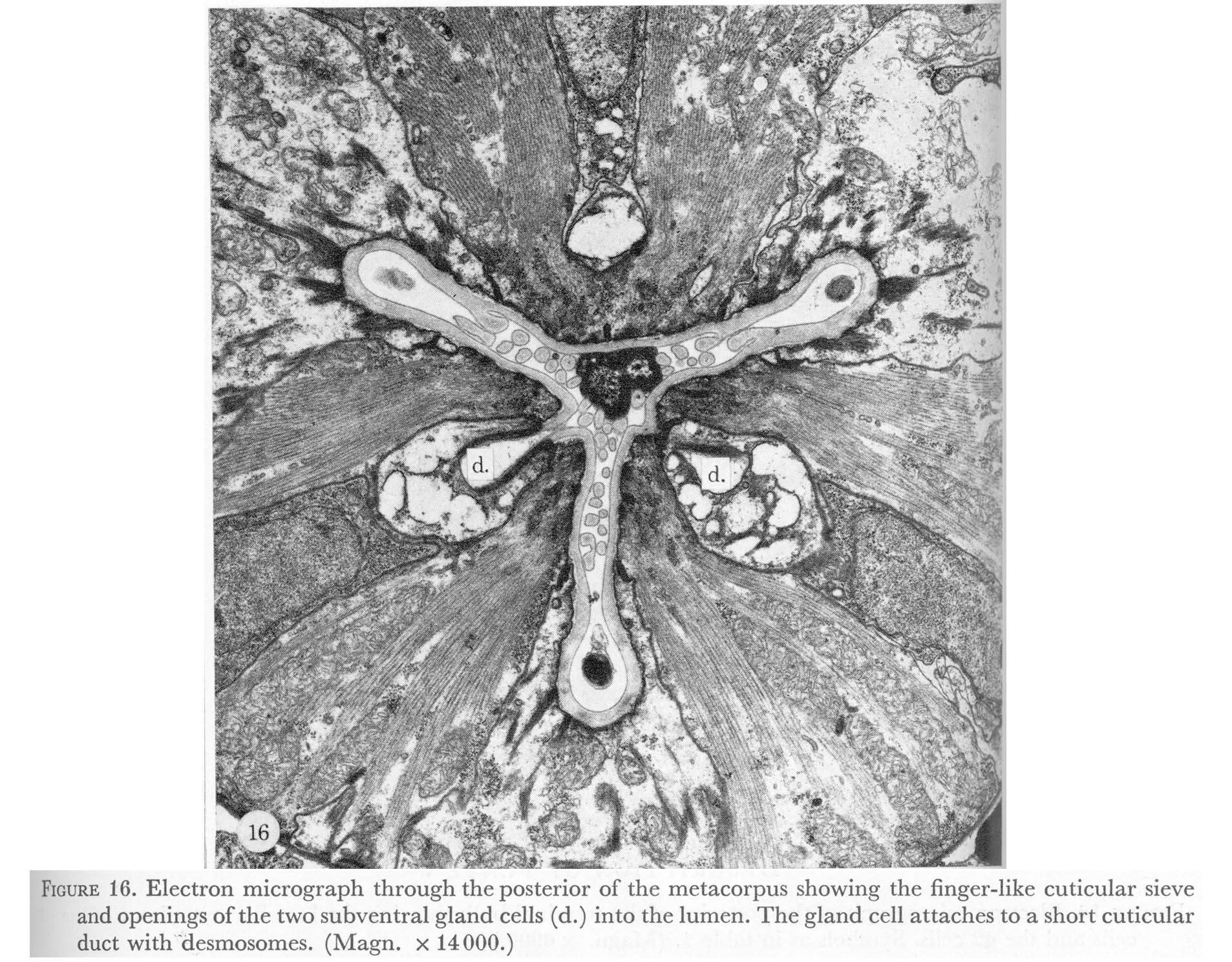
The two other regions of cuticle specialization appear to be for trapping and grinding food. Between the metacorpus and isthmus fingers of cuticle extend posteriorly in the pharyngeal lumen for 20 um when the pharynx is at rest. Upon contraction these fingers probably are pulled forward and mesh to form a sieve, which could trap particulate food matter, while excess liquid is expelled as the pharyngeal muscles relax. Possibly, it also mixes the contents of the lumen with digestive enzymes from the subventral gland cells which open into the lumen here. In figure 16, plate 6, bacteria trapped in this structure at the time of fixation can be seen.
A T-shaped region in the middle of the terminal bulb in figure 1, plate 1 marks the differentiation of the cuticle into the knobbed grinding organ shown in the electron micrograph in figure 17, plate 6. Muscle fibres run both radially and longitudinally here so that when these cells contract the cuticle knobs will not only be brought into apposition, but might also be moved forwards and backwards to promote mixing and grinding of material in the lumen. The pair of ventral gland cells opens into the lumen slightly anterior to this specialization.
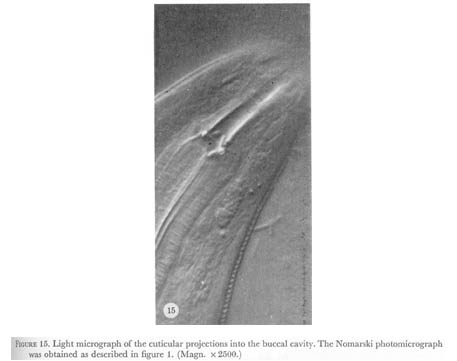
Figure 15. Plate 6 - Light micrograph of the cuticular projections into the buccal cavity. The Nomarski photomicrograph was obtained as described in figure 1. (Magn. X 2500.)
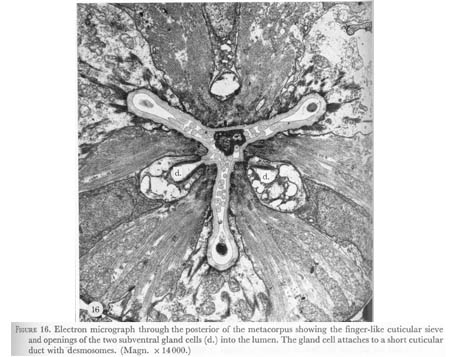 Figure 16. Plate 6 - Electron micrograph through the posterior of the metacorpus showing the finger-like cuticular sieve and openings of the two subventral gland cells (d.) into the lumen. The gland cell attaches to a short cuticular duct with desmosomes. (Magn. x 14000.)
Figure 16. Plate 6 - Electron micrograph through the posterior of the metacorpus showing the finger-like cuticular sieve and openings of the two subventral gland cells (d.) into the lumen. The gland cell attaches to a short cuticular duct with desmosomes. (Magn. x 14000.)
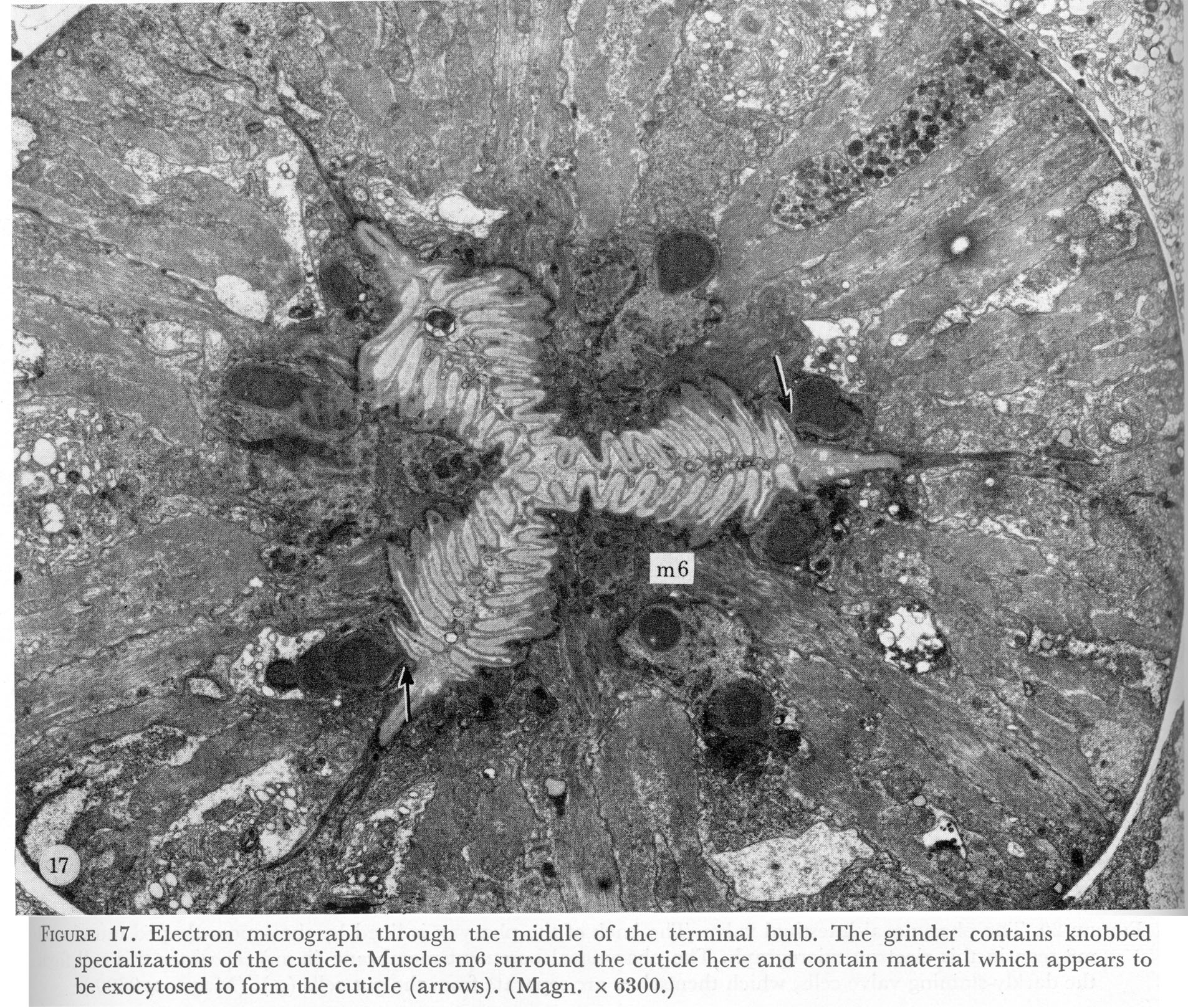
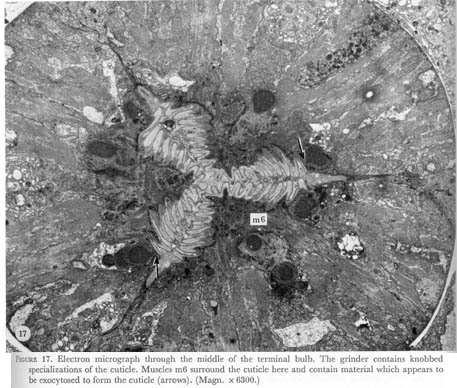 Figure 17. Plate 6 - Electron micrograph through the middle of the terminal bulb. The grinder contains knobbed specializations of the cuticle. Muscles m6 surround the cuticle here and contain material which appears to be exocytosed to form the cuticle (arrows). (Magn. x 6300.)
Figure 17. Plate 6 - Electron micrograph through the middle of the terminal bulb. The grinder contains knobbed specializations of the cuticle. Muscles m6 surround the cuticle here and contain material which appears to be exocytosed to form the cuticle (arrows). (Magn. x 6300.)
(c) Marginal cells
At the apices of the lumen the marginal cells are joined to the adjacent muscle cells by desmosomes. Darkly-staining filaments extend from the pharyngeal basement membrane to the cuticle (figures 5 and 6, plates 2 and 3). There are a total of nine marginal cells, the first three extend from the most anterior muscle cell in the procorpus to the level of the nerve ring in the metacorpus, the second set from here to the anterior end of the terminal bulb. The last set of three cells in the terminal bulb surround the pharyngeal cuticle at the posterior part and are syncytial (figure 18, plate 7).
The function of the marginal cells has occasioned much debate. Positioned at the apices of the lumen, a structural role seemed most likely to some authors. Others, seeing the darkly-staining filaments, thought them to be contractile (for a review see de Coninck 1965). In C. elegans they may well be contractile, since they receive innervation from the pharyngeal nervous system. A pair of marginal cell neurones synapse only on the second set of marginal cells, while axon terminals from motor neurone M5 ramify deeply into the third set of marginal cells near the cuticle, and both the muscles of the terminal bulb and the third set of marginal cells appear to be possible post-synaptic cells. Nervous innervation of the anterior set of marginal cells is weak in comparison. Motor neurones M2 which send a commissure dorsally from the subventral nerve cords occasionally synapse on the muscle cells here and on the subdorsal two marginal cells. The ventral marginal cell is not innervated, since these neurones do not cross the ventral side.
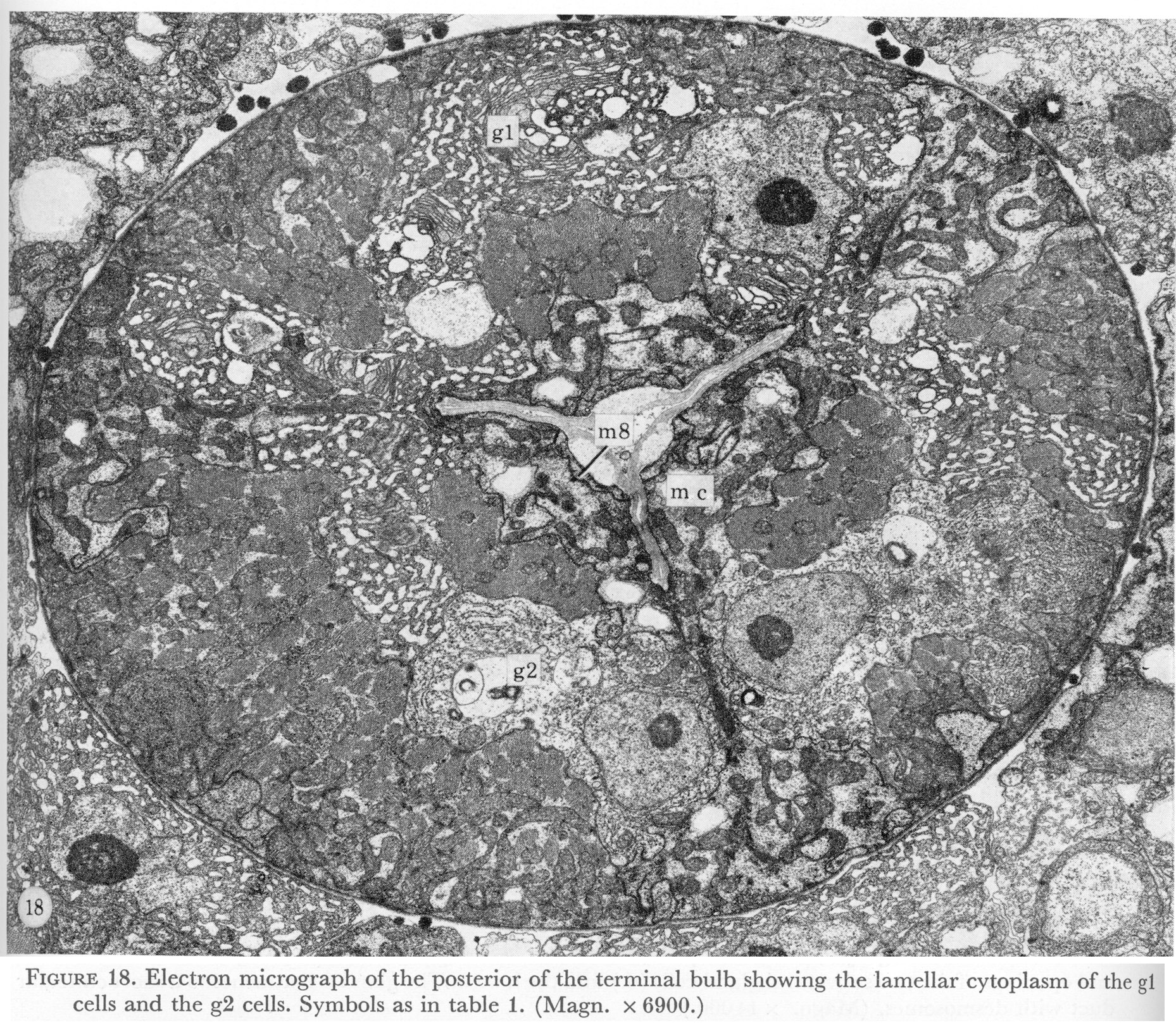
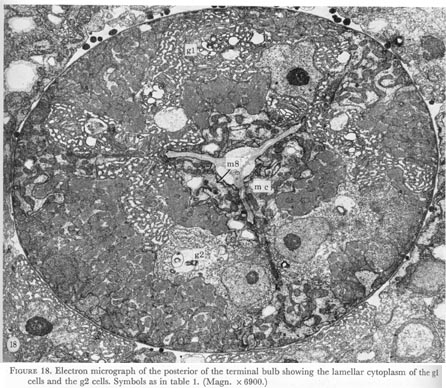 Figure 18. Plate 7 - Electron micrograph of the posterior of the terminal bulb showing the lamellar cytoplasm of the g1 cells and the g2 cells. Symbols as in table 1. (Magn. x 6900.)
Figure 18. Plate 7 - Electron micrograph of the posterior of the terminal bulb showing the lamellar cytoplasm of the g1 cells and the g2 cells. Symbols as in table 1. (Magn. x 6900.)
(d) The gland cells
Two types of gland cells may be distinguished by the fine structure of the cytoplasm. A set of three cells, called g1, have a lamellar cytoplasm, while a ventral pair of cells, g2, are clearer and vesicles can be seen in them. A section through the posterior part of the terminal bulb (figure 18, plate 7) shows the ventral g2 cells and the dorsal g1 cell. Both types of gland cell open into the lumen via a short cuticular duct, and are attached to it by means of desmosomes (figure 16, plate 6).
In the terminal bulb gland cell bodies swell and occupy much of this region of the pharynx (figure 19). Here also the dorsal and right g1 cells fuse. The gland cells may receive innervation from motor neurones M4 and M5 which synapse on muscle and gland cells together. In addition, the cell bodies of motor neurones M2 attach to the gland cells by desmosome-like junctions (figure 11, plate 5).
The function of these glands cannot be assigned from the anatomy, but their possible coordinate innervation with muscle cells suggests that they might be stimulated to secrete digestive enzymes when the isthmus and terminal bulb of the pharynx open. Movements of vesicles within the g1 cells are visible during moulting when the posterior two-thirds of the pharyngeal cuticle is shredded and ingested by contractions of the terminal bulb (J. Sulston, personal communication). Hence secretions from these glands may aid in this degradative process.
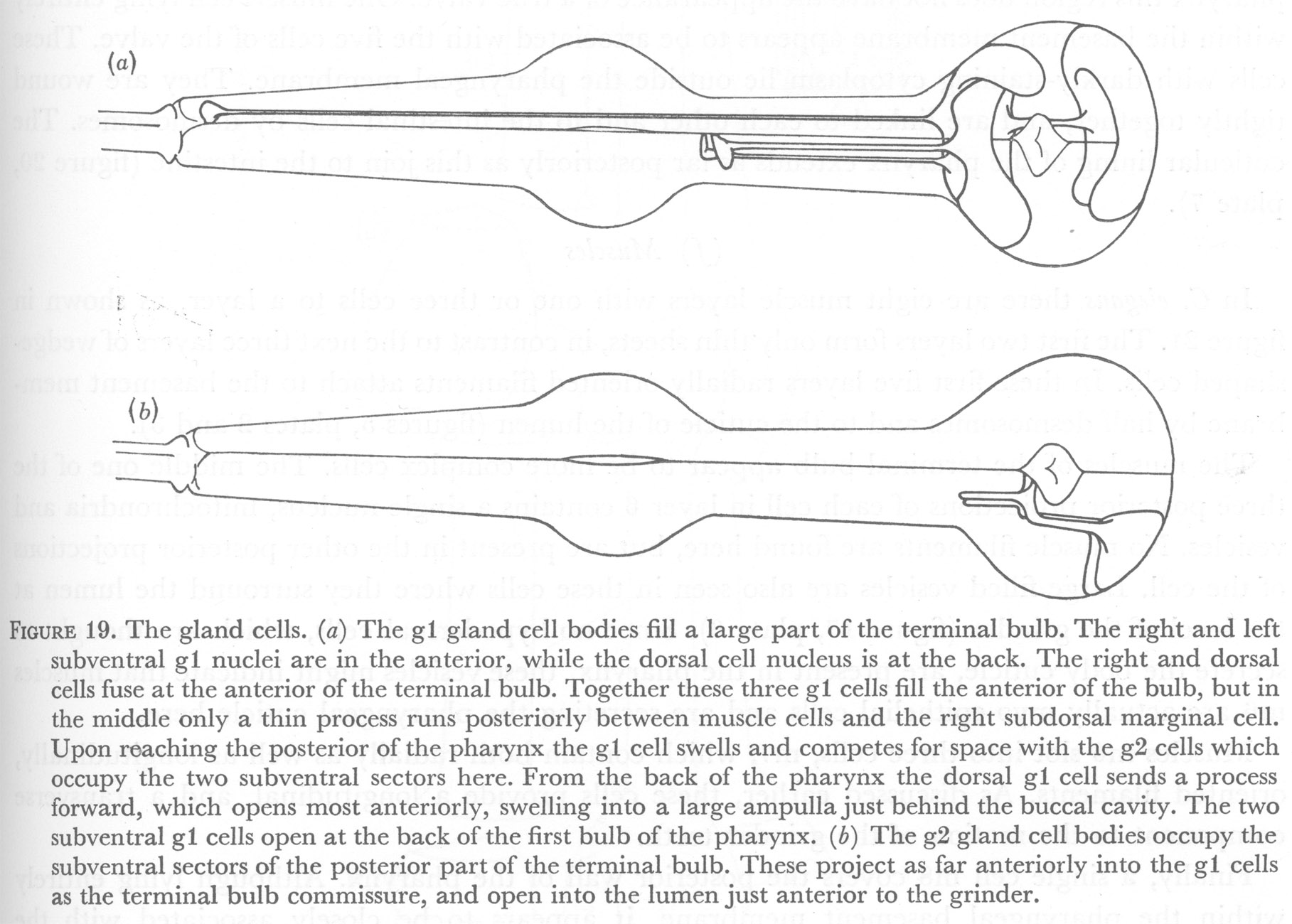
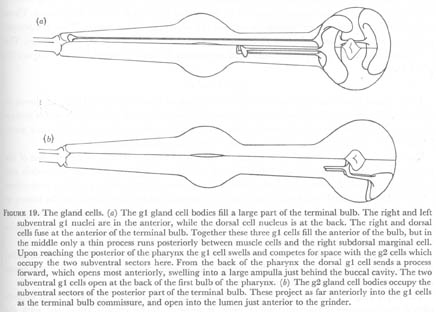 Figure 19. The gland cells. (a) The g1 gland cell bodies fill a large part of the terminal bulb. The right and left subventral g1 nuclei are in the anterior, while the dorsal cell nucleus is at the back. The right and dorsal cells fuse at the anterior of the terminal bulb. Together these three g1 cells fill the anterior of the bulb, but in the middle only a thin process runs posteriorly between muscle cells and the right subdorsal marginal cell. Upon reaching the posterior of the pharynx the g1 cell swells and competes for space with the g2 cells which occupy the two subventral sectors here. From the back of the pharynx the dorsal g1 cell sends a process forward, which opens most anteriorly, swelling into a large ampulla just behind the buccal cavity. The two subventral g1 cells open at the back of the first bulb of the pharynx. (b) The g2 gland cell bodies occupy the subventral sectors of the posterior part of the terminal bulb. These project as far anteriorly into the g1 cells as the terminal bulb commissure, and open into the lumen just anterior to the grinder.
Figure 19. The gland cells. (a) The g1 gland cell bodies fill a large part of the terminal bulb. The right and left subventral g1 nuclei are in the anterior, while the dorsal cell nucleus is at the back. The right and dorsal cells fuse at the anterior of the terminal bulb. Together these three g1 cells fill the anterior of the bulb, but in the middle only a thin process runs posteriorly between muscle cells and the right subdorsal marginal cell. Upon reaching the posterior of the pharynx the g1 cell swells and competes for space with the g2 cells which occupy the two subventral sectors here. From the back of the pharynx the dorsal g1 cell sends a process forward, which opens most anteriorly, swelling into a large ampulla just behind the buccal cavity. The two subventral g1 cells open at the back of the first bulb of the pharynx. (b) The g2 gland cell bodies occupy the subventral sectors of the posterior part of the terminal bulb. These project as far anteriorly into the g1 cells as the terminal bulb commissure, and open into the lumen just anterior to the grinder.
(e) Pharyngeal-intestinal valve
The lumen of the pharyngeal-intestinal valve is triangular and open, so that in the resting pharynx this region does not have the appearance of a true valve. One muscle cell lying entirely within the basement membrane appears to be associated with the five cells of the valve. These cells with darkly-staining cytoplasm lie outside the pharyngeal membrane. They are wound tightly together, and are linked to each other and to the intestinal cells by desmosomes. The cuticular lining of the pharynx extends as far posteriorly as this join to the intestine (figure 20, plate 7).
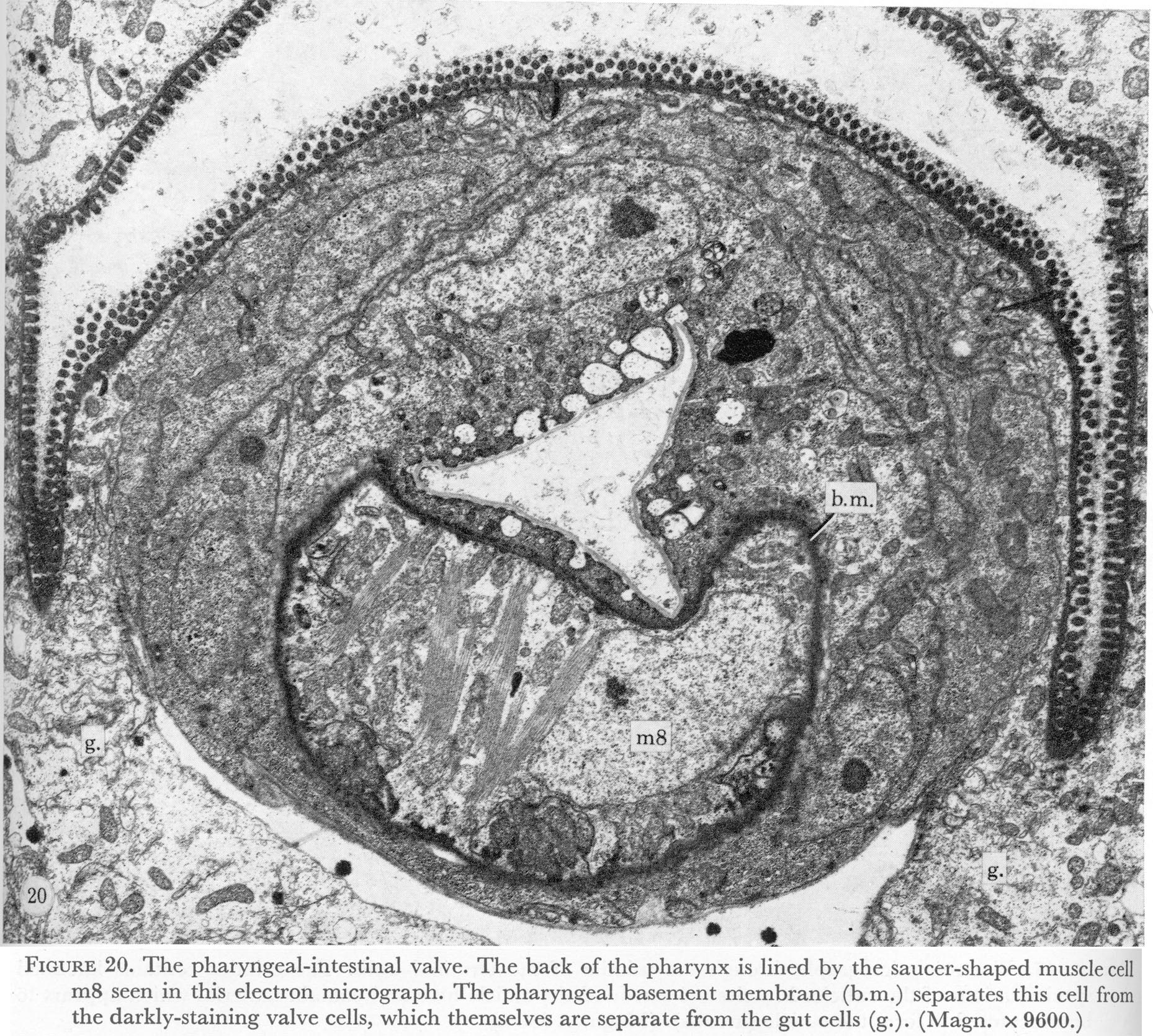
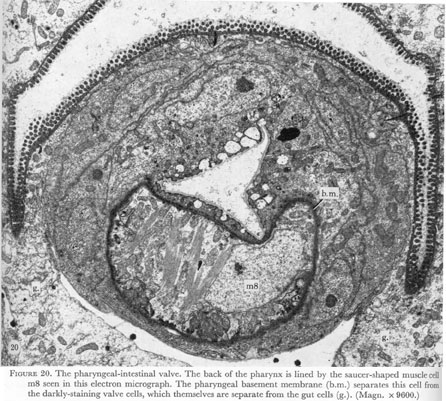 Figure 20. Plate 7 - The pharyngeal-intestinal valve. The back of the pharynx is lined by the saucer-shaped muscle cell m8 seen in this electron micrograph. The pharyngeal basement membrane (b.m.) separates this cell from the darkly-staining valve cells, which themselves are separate from the gut cells (g.). (Magn. X 9600.)
Figure 20. Plate 7 - The pharyngeal-intestinal valve. The back of the pharynx is lined by the saucer-shaped muscle cell m8 seen in this electron micrograph. The pharyngeal basement membrane (b.m.) separates this cell from the darkly-staining valve cells, which themselves are separate from the gut cells (g.). (Magn. X 9600.)
(f) Muscles
In C. elegans there are eight muscle layers with one or three cells to a layer, as shown in figure 21. The first two layers form only thin sheets, in contrast to the next three layers of wedge-shaped cells. In these first five layers radially oriented filaments attach to the basement membrane by half desmosomes and to the cuticle of the lumen (figures 5, plates 2 and 3).
The muscles of the terminal bulb appear to be more complex cells. The middle one of the three posterior projections of each cell in layer 6 contains a single nucleus, mitochondria and vesicles. No muscle filaments are found here, but are present in the other posterior projections of the cell. Large filled vesicles are also seen in these cells where they surround the lumen at the level of the grinder (figure 17, plate 6). Since no hypodermal cells, which are thought to secrete the body cuticle, are present in the pharynx, these vesicles might indicate that muscles m6 are actually myo-epithelial cells and are secreting the pharyngeal cuticle here.
Muscles m6 slot into three cells, m7, which contain both radially as well as longitudinally, oriented filaments. As discussed earlier, these cells provide a longitudinal, and a transverse component to the motion of the grinder teeth.
Finally, a single cell m8 covers the posterior wall of the pharynx. Although lying entirely within the pharyngeal basement membrane, it appears to be closely associated with the pharyngeal-intestinal valve (figure 20, plate 7). The most anterior projections of m8 lie next to the pharyngeal cuticle completely surrounded on the outside by the syncitial marginal cells 3 (figure 18, plate 7). Thus the cell is completely sequestered from the most posterior motor neurone, M5 and no neuromuscular junctions with m8 can be found. However m8 is joined to the marginal cells 3 by desmosomes and might then receive innervation by gap junctions to the marginal cells, which themselves receive chemical synapses from M5.
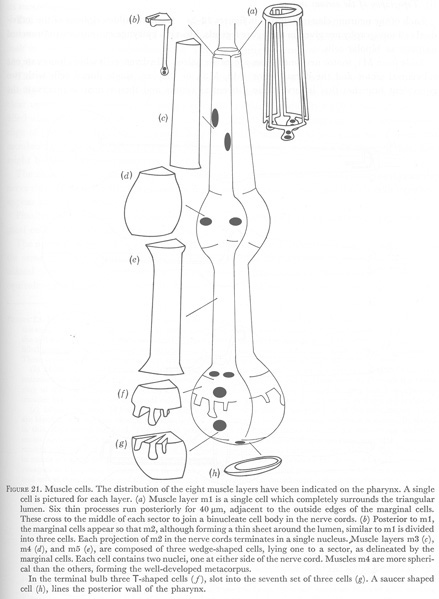 Figure 21. Muscle cells. The distribution of the eight muscle layers have been indicated on the pharynx. A single cell is pictured for each layer. (a) Muscle layer m1 is a single cell which completely surrounds the triangular lumen. Six thin processes run posteriorly for 40 um, adjacent to the outside edges of the marginal cells. These cross to the middle of each sector to join a binucleate cell body in the nerve cords. (b) Posterior to m1, the marginal cells appear so that m2, although forming a thin sheet around the lumen, similar to m1 is divided into three cells. Each projection of m2 in the nerve cords terminates in a soma with two nuclei. Muscle layers m3 (c), m4 (d), and m5 (e), are composed of three wedge-shaped cells, lying one to a sector, as delineated by the marginal cells. Each cell contains two nuclei, one at either side of the nerve cord. Muscles m4 are more spherical than the others, forming the well-developed metacorpus.
Figure 21. Muscle cells. The distribution of the eight muscle layers have been indicated on the pharynx. A single cell is pictured for each layer. (a) Muscle layer m1 is a single cell which completely surrounds the triangular lumen. Six thin processes run posteriorly for 40 um, adjacent to the outside edges of the marginal cells. These cross to the middle of each sector to join a binucleate cell body in the nerve cords. (b) Posterior to m1, the marginal cells appear so that m2, although forming a thin sheet around the lumen, similar to m1 is divided into three cells. Each projection of m2 in the nerve cords terminates in a soma with two nuclei. Muscle layers m3 (c), m4 (d), and m5 (e), are composed of three wedge-shaped cells, lying one to a sector, as delineated by the marginal cells. Each cell contains two nuclei, one at either side of the nerve cord. Muscles m4 are more spherical than the others, forming the well-developed metacorpus.
In the terminal bulb three T-shaped cells (f), slot into the seventh set of three cells (g). A saucer shaped cell (h), lines the posterior wall of the pharynx.
WA editors' note: In the original publication this image was partly mirror-reversed. Corrected version is shown here (L. Avery pers. comm.). The nuclei (black spheres) have been added to the image by WA. The number of nuclei for m2 has been corrected as two. Also see corrected image for pharyngeal muscles in C. elegans Book II and a colored version, PhaFIG 6, in the Pharynx section of the Handbook.
(g) Nervous system
The nervous system of the pharynx is composed of 20 cells (table 1). In classifying these cells it became apparent that frequently pairs of bilaterally symmetric cells were functionally identical. These could then be treated as a single group of cells, and five classes of motor neurones (7 cells) and six classes of interneurones (8 cells) were identified. The remaining five cells were divided into three types which did not fall into either of these categories and were classed simply as other neurones.
(i) Topography of the nervous system
Each of the neurone classes is drawn in figures 22-24, and detailed descriptions of the individual cell topography are given in the figure legends. Most pharyngeal neurones are unbranched unipolar or bipolar cells. Aside from M1, motor neurone classes are either paired subventral cells which innervate one subventral sector and the dorsal sector (M2, M3), or they are single dorsal cells with two equivalent branches that innervate the subventral sectors and then return to innervate the dorsal sector (M4, M5). The first motor neurone, M1 is a dorsal cell that branches as it makes its motor synapses.
The three interneurone classes of the corpus (I1-I3) are unbranched bipolar cells, as is I6. Interneurone I5 is a complex cell that closes with itself in the nerve ring and is again joined to itself in the terminal bulb commissure. The remaining interneurone, I4, similar to M4 and M5, is a single dorsal cell with two equivalent branches.
The pair of neurosecretory-motor neurones drawn in figure 24a are bipolar cells which contain lightly- and darkly-staining membrane bound vesicles 200 nm in diameter, as well as small clear vesicles. In the isthmus, branches from these cells periodically swell to form varicosities filled with both types of vesicle (figures 7 and 8, plate 3). The processes run at the most exterior edge of the nerve cords and synaptic bars are found adjacent to the pharyngeal basement membrane, or muscle cells (figure 7). Thus, the morphology of these cells suggests that they might be both neurosecretory and motor. The motor-interneurone (figure 24b) is a single unipolar dorsal cell that circumnavigates the nerve ring. In so doing weak synapses are made, and often both nerve and muscle cells together, appear to be post-synaptic.
Finally, the marginal cell neurones are a pair of bipolar cells which innervate only the marginal cells in the anterior of the isthmus.
The nerve ring formed by these cell processes is itself a well-organized structure, and appears the same in different animals. This can be seen by selecting a portion of the nerve ring in one animal and tracing the cell sections in each sector (figure 25a-c), then comparing these to equivalent regions in other animals (figure 25d-f). Looking at the nerve ring in this way, one sees that (1) pairs of subventral cells (or branches of M4, M5, I4) occupy equivalent positions relative to other cells on the right and left side of the ring (compare positions of I2 and NSM, for example), (2) pairs of subventral cells (or branches of M4, M5, I4) which cross over the dorso-ventral midline exchange positions relative to each other (compare MC neurones in figure 25a) and (3) pairs of cells or branches of a single cell that terminate on the dorsal side meet in gap junctions on the dorsal midline (figure 12). Besides these characteristics some cells are closely associated with others and are very noticeable features of the nerve ring. For example, I5 sits on top of M4, I2 are adjacent to NSM, and MI grows clockwise around the ring, associated with the branches of I4. These observations on the organization of the neuropil suggest that cell growth is not random here, but that well defined paths are laid out for cells to form this structure.
(ii) Connectivity
Pharyngeal neurones make synapses en passant similar to the ventral cord neurones (White et al. 1976). Therefore no 'synaptic endings' or 'synaptic terminals' in the generally accepted sense are present, and in the following discussion 'ending' and 'termination' simply refer to the most distal tip of the neuronal process.
A. Input to the pharyngeal nervous system
Two possible inputs can be distinguished. One is external, from the somatic nervous system, and the other, internal sources of input might be the endings of some pharyngeal neurones themselves. As will be described below these appear to be proprioreceptors or mechanoreceptors.
(1) Somatic pharyngeal innervation. Only a single pair of interneurones that receives synapses in the central nerve ring cross the basement membrane of the pharynx (Ward et al. 1975). These enter the subventral nerve cords where they begin at the first muscle layer.
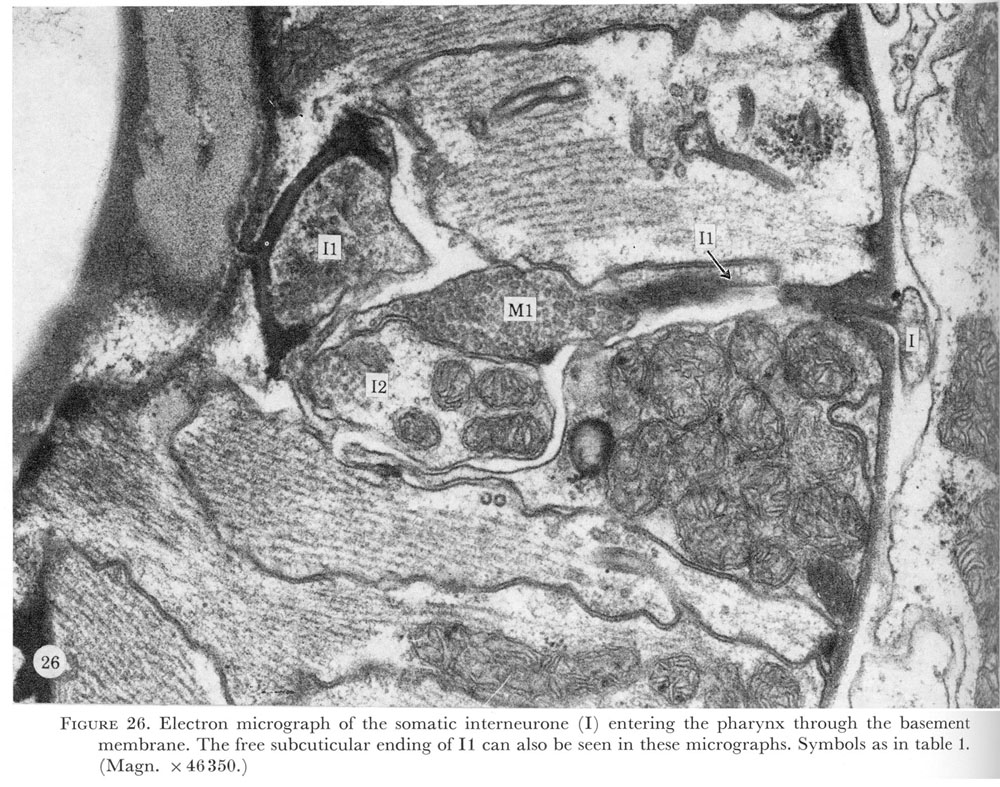
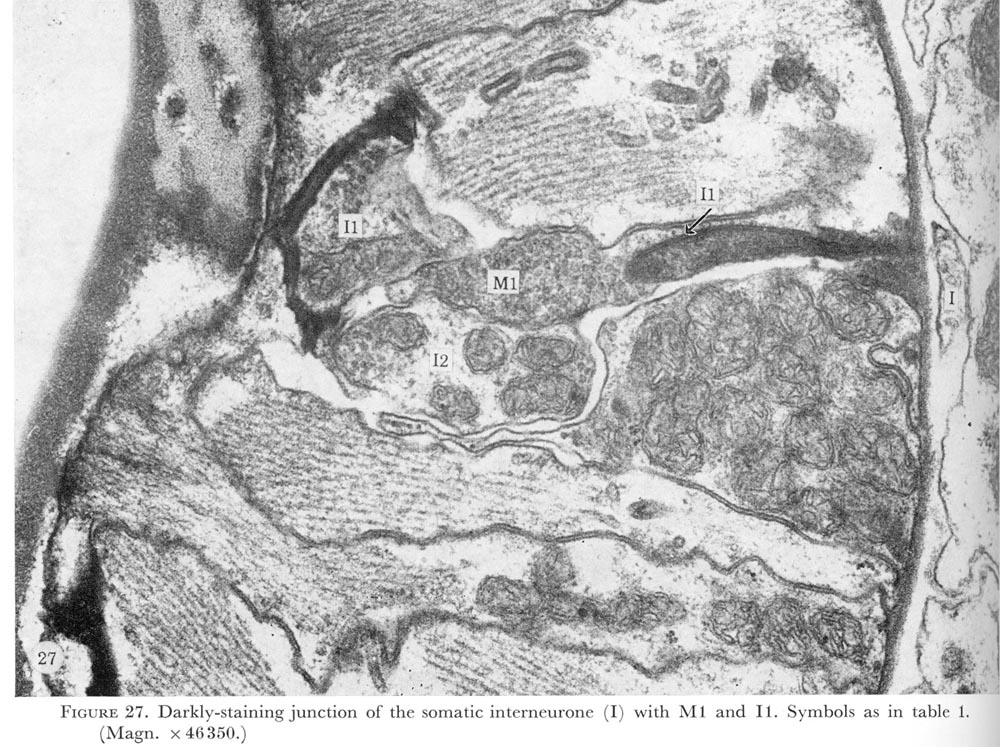
Just before the neurone passes through the basement membrane a darkly-staining structure in the end of the neurone can be seen (figure 26, plate 8). The ending of the interneurone swells and is buried in the pharyngeal interneurone I1 (figure 27, plate 8). The pharyngeal motor neurone M1 is also associated with this unusual neuronal ending. As indicated in figure 3 possibly both of these neurones make electrical connections with the ring interneurones.
(2) Mechano-reception. Several neurone cell classes listed in table 1 have free endings just under the cuticle of the pharyngeal lumen. The neurone is fastened to the adjacent cell or cells by desmosomes, and when the position of the lumen changes these cells might fire because of deformation of their endings. If so, then these cells could be mechanical sensory receptors, and for convenience have been called proprio- or mechanoreceptive when describing the presence of such endings in various neurone classes.
Two types of ending have been distinguished on the basis of the morphology of the adjacent cells. Endings of interneurones are similar to that of I1 shown in figures 26 and 27, plate 8. In contrast, the motor neurones M3, the neurosecretory motor neurones, and the marginal cell neurones all attach to muscle cells in which the cytoplasm contains darkly-staining striations, as shown in figure 10, plate 4. Interneurone I5 attaches to the cell bodies of M2 and here the cytoplasm of the neurone cell body is reticulated (figure 13, plate 5) and appears similar to the muscle cell cytoplasm associated with the other neuronal endings. Gland cells occupy much of this region of the terminal bulb, and if these are easily deformed by muscle contractions, then I5 could also be a type of mechanoreceptor.
B. Pharyngeal connectivity
Using the criteria described by White et al. (1976) gap junctions and chemical synapses were assigned for cells on as many animals as there were reliable series, or portions of series. In all cases at least two animals could be compared. In the pharynx synaptic structures generally extend over at least two sections or micrographs. Figures 28-30 represent a compilation of all the available data for each cell. These connections were then summarized in the two diagrams in figure 32. The reasons for separating these two circuits will be taken up in the discussion.
(1) Motor innervation. Motor neurone M1 couples the first three muscle layers together to innervate the procorpus. It does this by sending two branches from the dorsal nerve cord to make synapses on muscles m1 and m2 en passant to the subventral cords, while synapses to m3 are made from the three nerve cords (figure 28).
The pair of motor neurones M2 synapse on muscles m5 from the subventral nerve cords, and to muscles m4 anterior to the nerve ring as these cells cross to the dorsal midline (figure 28). Muscles m4 are also driven by a pair of cells M3, which does not make synapses to any other muscle cell.
In the isthmus a single cell M4 synapses to muscles m5 from all three nerve cords. Darkly-staining vesicles, similar to the 65 nm dense core vesicles associated with some amphidial neurones (Ware et al. 1975; J. G. White, personal communication) are seen in this cell which are not seen in other motor neurones of the pharynx (compare motor neurones M4 and M5 in figure 11, plate 5). All other pharyngeal neurones contain the clear 40 nm vesicles described by Ware et al. (1975) in the nerve ring of C. elegans.
Both muscles m6 and m7 in the terminal bulb are innervated by a single motor neurone M5.
(2) Interneurones. The connections formed by interneurones I1 are summarized in figure 29a. At the anterior of the procorpus the endings of the neurones dip under the cuticle of the lumen to form possible proprioreceptive endings, and they also reach out to the basement membrane here to receive input from the somatic nervous system. Anterior to the ring these cells synapse on M2 and M3 on their respective subventral sides, and again on the dorsal side to the same member of these paired motor neurones or the partner from the opposite side. In a similar fashion, synapses are made to the NSM neurones, and gap junctions are made to the marginal cell neurones. Some weak synapses to I3 and I5 have also been found, but this is variable. At the most anterior end of the subventral nerve cords interneurones I2 have a proprioreceptive-like ending and they are coupled by gap junctions to M1. Some chemical synapses are made to M1 here, but M1 also makes synapses in which both I2 and m3 appear to be the post-synaptic cells. These connections vary between animals. Sometimes M1 synapses on both I2 cells, sometimes on one and sometimes no synapses from M1 to I2 are found. In the nerve ring one or both I2 cells synapse characteristically on M1 near the right subdorsal apex of the lumen (figures 28a and 29b). Also characteristic of I2 in the nerve ring is their association with the NSM cells. These chemical synapses, which have relatively larger post-synaptic darkenings and many vesicles, are made to the NSM cells on the subventral sides anterior to the ring. In the ring I2 cells are full of vesicles and form synapses on the NSM cells which extend over five or more sections, while synapses on I6 on the dorsal side of the nerve ring can be found on only one or two sections. After crossing the dorsal side I2 run posteriorly in the opposite subventral nerve cord now synapsing on the other member of the NSM pair and on I4 (figure 29b).
Interneurone I3 has a proprioreceptive-like ending at the beginning of the dorsal nerve cord. Here also, I3 and m3 may both be post-synaptic cells, receiving synapses from M1. Neurone I3 ends on the dorsal side of the nerve ring and forms one or two synapses on M3 and to one or the other member of the pair M2. Synapses are also made to I6 and the NSM cells.
Most characteristic of I4 are chemical synapses on the cell bodies of M3 which may continue through five to ten sections. Synapses are also made on the projections of M3 near the cell body and frequently these appear to be made on the NSM cell as well. Interneurone I4 receives from I2 in this same region. In the ring I4 makes further synapses to M3 and receives from I2, but no connections to NSM cells are made here, suggesting that connections to M3 and from I2 are its strongest features.
Similarly, I5 makes some synapses in which M3 and M4 appear to be post-synaptic cells. Its association with M4 however is much stronger and it sits on top of this cell in the nerve ring. A single synapse to M1 is made in the ring. Either chemical synapses can be received from I1, or I1 and I5 can be coupled by gap junctions. In the terminal bulb gap junctions are made to M5.
At the level of the terminal bulb commissure I6 has a proprioreceptive ending and makes gap junctions to M5. In the metacorpus I6 synapses on M4 and receives from 12.
(3) Other neurones.The neurosecretory motor neurones receive very extensive synapses from I2, as discussed previously. Fewer and smaller synapses from I1 and I3 are also seen. In the anterior of the isthmus I6 synapses on the NSM cells, but these also appear to synapse on muscles and 16 more posteriorly. The NSM cells are proprioreceptive at the level of the nerve ring.
The motor-interneurone makes synapses that often have both nerve and muscle as the post synaptic cells. Its connections are variable and generally chemical synapses or gap junctions are made to M1, M2, M3, I3, I4, I5 and the marginal cell neurones.
The marginal cell neurones are proprioreceptive at the back of the procorpus. Their most prominent connections are gap junctions to M2, but they also receive some chemical synapses from I1 and make gap junctions with I1 (figure 30c). They synapse strongly on the marginal cells 2.
(iii) Variability
In the diagrams in figures 28-30 strong synaptic features of cells appear to be those that are repeated at one or more places on the cell. For example, pairs of subventral cells almost always synapse on a target cell on the ipsilateral side, as well as to it and/or its partner on the dorsal side. Synapses that are least reproducible from animal to animal and are therefore more variable are those in which there appear to be two post-synaptic cells. The motor-interneurone is the most extreme example of this. Two cells are shown in figure 30, because common features are not readily apparent. As mentioned in the preceding sections, in some cases a connection between two cells might be made by gap junctions in one animal and chemical synapses in another. Variations that were seen in different animals are indicated in parentheses in figures 28-30. Often synapses on the dorsal side of the nerve ring appear more sloppy.
The terminations of the I1 cells are variable, although their connections are not. Generally, one of the I1 cells continues anteriorly in the dorsal nerve cord after reaching the dorsal side of the nerve ring. Its partner might do the same for a short distance, or might continue around the nerve ring to end on the opposite subventral side.
Aside from variable endings, the remarkable constancy of cell morphology in the nematode is an aid in the reconstruction of its nervous system. However, occasional gross alterations in branching patterns of cell processes do occur in wild type animals, which may suggest some of the mechanisms used for development of the nervous system. In a single wild type animal several differences were found in the nerve ring, and were compared with the two other animals with completely reconstructed nerve rings.
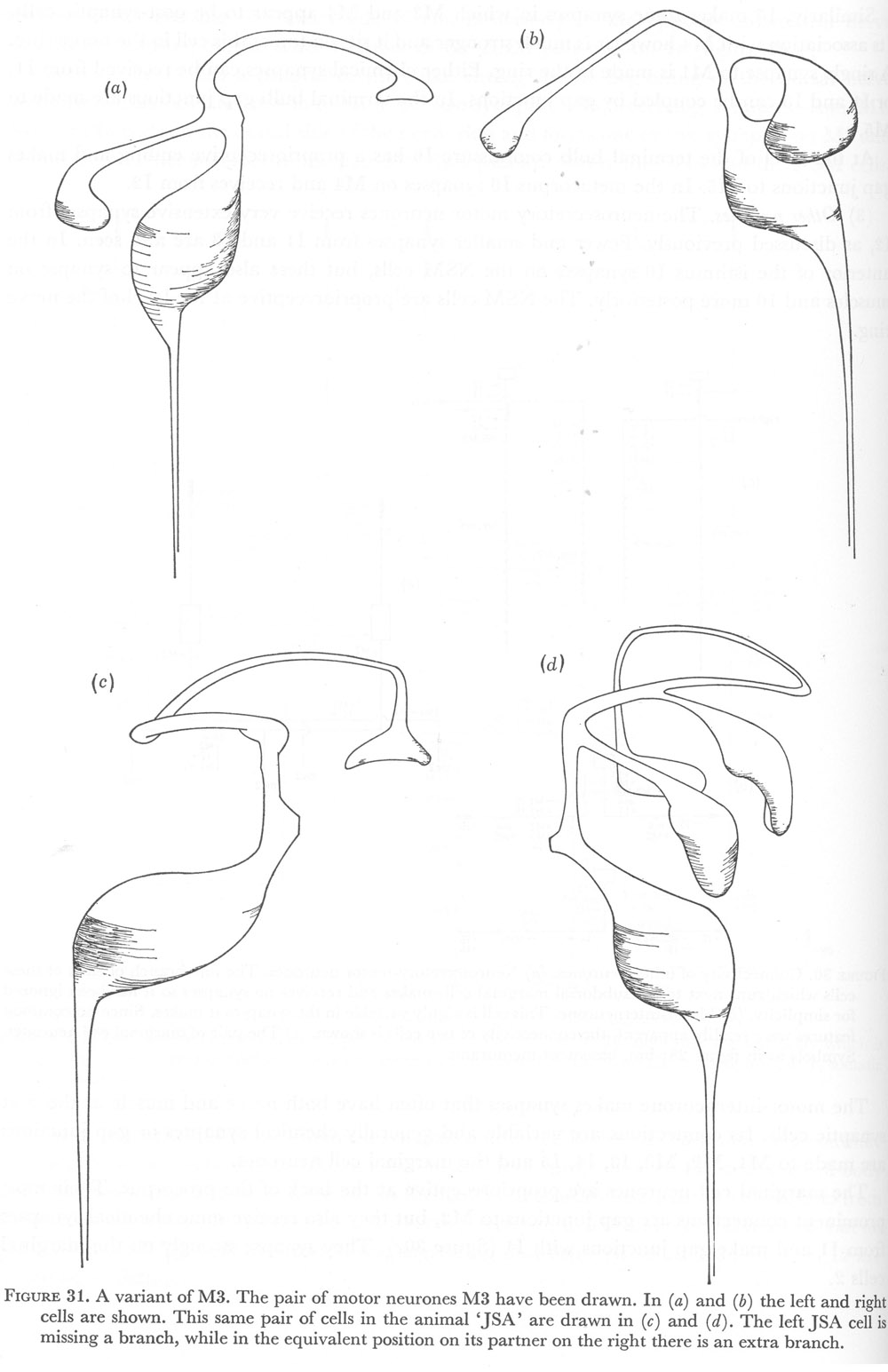
As illustrated in figure 31, the first alteration that was noticed was the absence of the anterior subventral branch of the left M3 cell. Synapses to muscles m4 on the subventral side were still made but at an unusual region of the cell. In contrast, the right partner had two of these subventral branches, and both branches formed synapses on muscles m4 in the customary manner. The remaining features of both cells were normal.
The destination of the posterior projection of the right marginal cell neurone was also unusual. After passing around the nerve ring to the left subventral sector this cell continued across the ventral midline and did not synapse on the ventral marginal cell. Rather, it crossed the right subventral sector and joined the left marginal cell neurone in synapsing on the right subdorsal marginal cell. It is interesting to note that where the cell crosses a region of the nerve ring it normally would not and so becomes an 'extra' process in this region, the cell still travels next to its characteristic set of cells. Figure 25e is a section through this region of the nerve ring which includes the extra marginal cell process.
Lastly, the motor-interneurone in this animal grew counterclockwise around the ring. Interneurone I4, which is closely associated with this cell in the ring sent its left branch, rather than the right branch to the dorsal side.
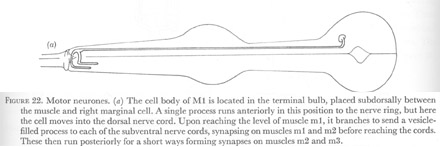
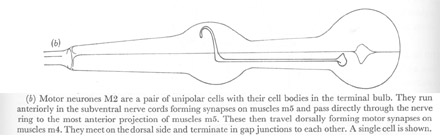
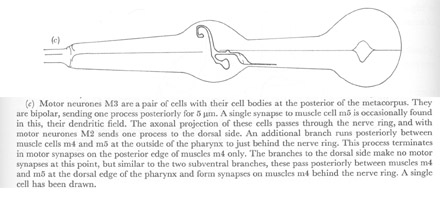
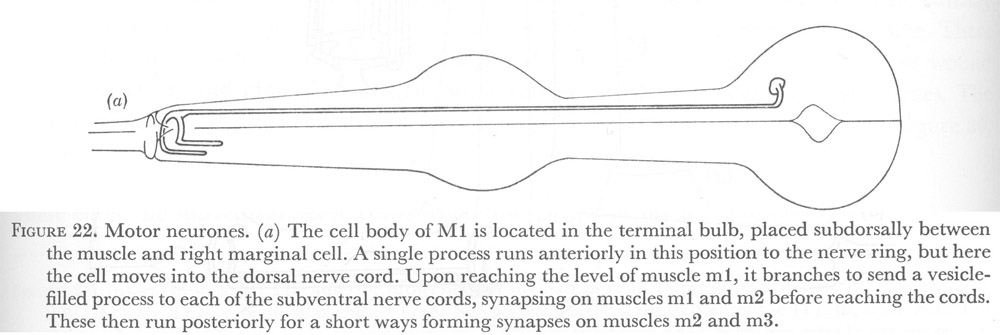
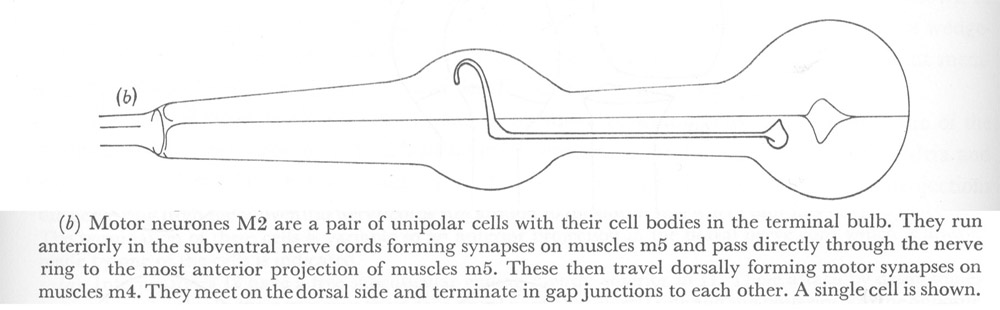
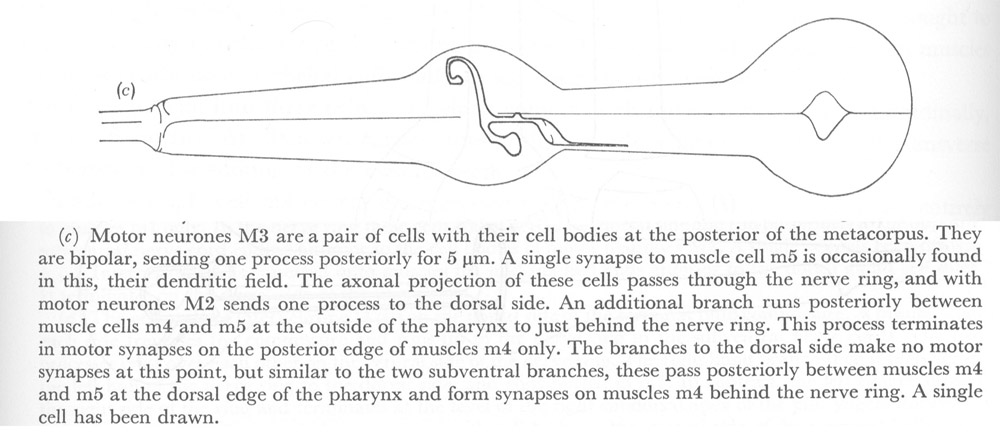
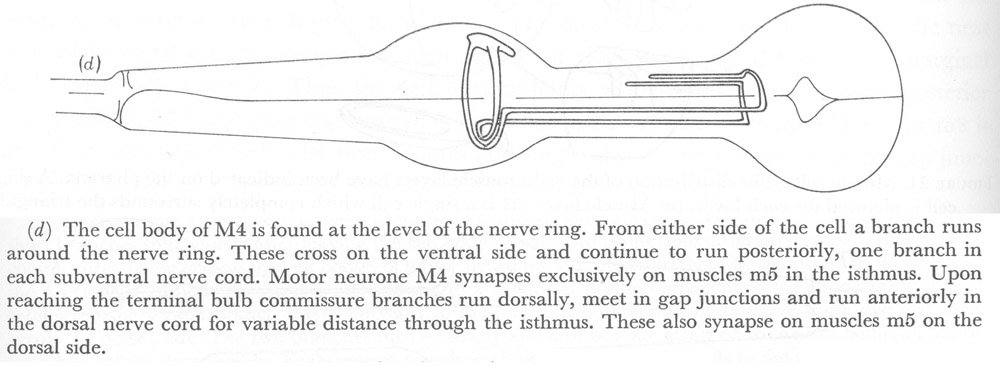
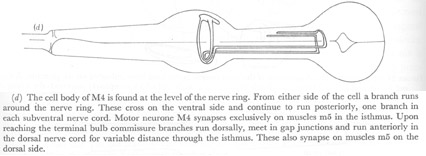 Figure 22. Motor neurones.
Figure 22. Motor neurones.
(a) The cell body of M1 is located in the terminal bulb, placed subdorsally between the muscle and right marginal cell. A single process runs anteriorly in this position to the nerve ring, but here the cell moves into the dorsal nerve cord. Upon reaching the level of muscle m1, it branches to send a vesicle-filled process to each of the subventral nerve cords, synapsing on muscles m1 and m2 before reaching the cords. These then run posteriorly for a short ways forming synapses on muscles m2 and m3.
(b) Motor neurones M2 are a pair of unipolar cells with their cell bodies in the terminal bulb. They run anteriorly in the sub ventral nerve cords forming synapses on muscles m5 and pass directly through the nerve ring to the most anterior projection of muscles m5. These then travel dorsally forming motor synapses on muscles m4. They meet on the dorsal side and terminate in gap junctions to each other. A single cell is shown.
(c) Motor neurones M3 are a pair of cells with their cell bodies at the posterior of the metacorpus. They are bipolar, sending one process posteriorly for 5 µm. A single synapse to muscle cell m5 is occasionally found in this, their dendritic field. The axonal projection of these cells passes through the nerve ring, and with motor neurones M2 sends one process to the dorsal side. An additional branch runs posteriorly between muscle cells m4 and m5 at the outside of the pharynx to just behind the nerve ring. This process terminates in motor synapses on the posterior edge of muscles m4 only. The branches to the dorsal side make no motor synapses at this point, but similar to the two subventral branches, these pass posteriorly between muscles m4 and m5 at the dorsal edge of the pharynx and form synapses on muscles m4 behind the nerve ring. A single cell has been drawn.
(d) The cell body of M4 is found at the level of the nerve ring. From either side of the cell a branch runs around the nerve ring. These cross on the ventral side and continue to run posteriorly, one branch in each subventral nerve cord. Motor neurone M4 synapses exclusively on muscles m5 in the isthmus. Upon reaching the terminal bulb commissure branches run dorsally, meet in gap junctions and run anteriorly in the dorsal nerve cord for variable distance through the isthmus. These also synapse on muscles m5 on the dorsal side.
(e) The cell body of M5 is on the left subdorsal, posterior to muscles m7. Like motor neurone M4 it branches at the cell body with one process running across the dorsal side behind muscles m7 and one runs ventrally. The two processes cross on the ventral side then run anteriorly adjacent to the ventral marginal cell to the terminal bulb commissure. The processes pass under the cell bodies of motor neurones M2 and split, sending a short arm posteriorly again on either side of the subdorsal marginal cells. These also meet in gap junctions on the dorsal side of the terminal bulb commissure.
WA editors' note: The actual locations of cell bodies of neurons in the metacorpus are more posterior to those shown here (L. Avery pers. comm.)
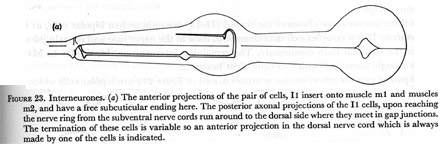
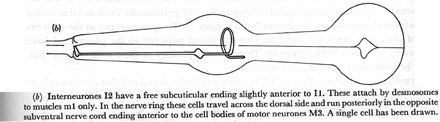
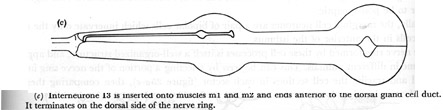
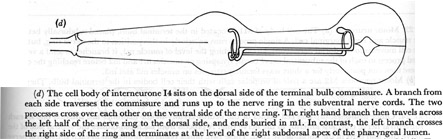
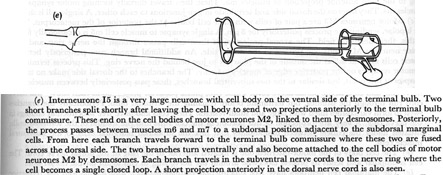
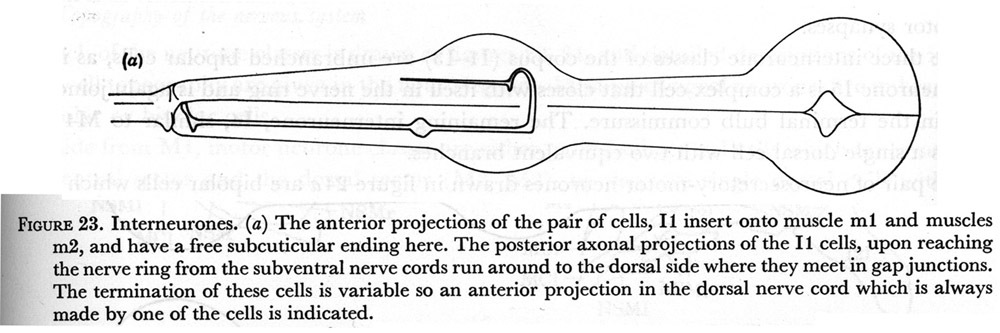
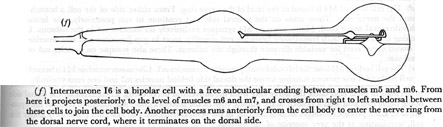 Figure 23. Interneurones.
Figure 23. Interneurones.
(a) The anterior projections of the pair of cells, I1 insert onto muscle m1 and muscles m2, and have a free subcuticular ending here. The posterior axonal projections of the I1 cells, upon reaching the nerve ring from the subventral nerve cords run around to the dorsal side where they meet in gap junctions. The termination of these cells is variable so an anterior projection in the dorsal nerve cord which is always made by one of the cells is indicated.
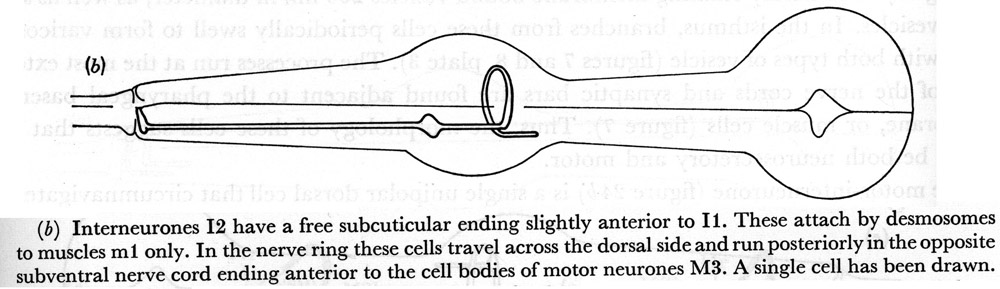
(b) Interneurones I2 have a free subcuticular ending slightly anterior to I1. These attach by desmosomes to muscles m1 only. In the nerve ring these cells travel across the dorsal side and run posteriorly in the opposite subventral nerve cord ending anterior to the cell bodies of motor neurones M3. A single cell has been drawn.
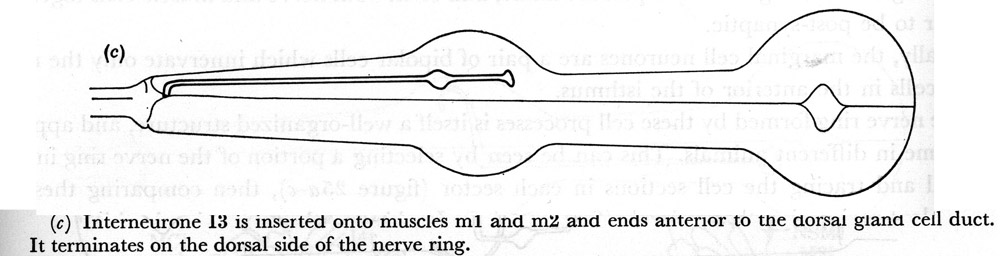
(c) Interneurone I3 is inserted onto muscles m1 and m2 and ends anterior to the dorsal gland cell duct. It terminates on the dorsal side of the nerve ring.
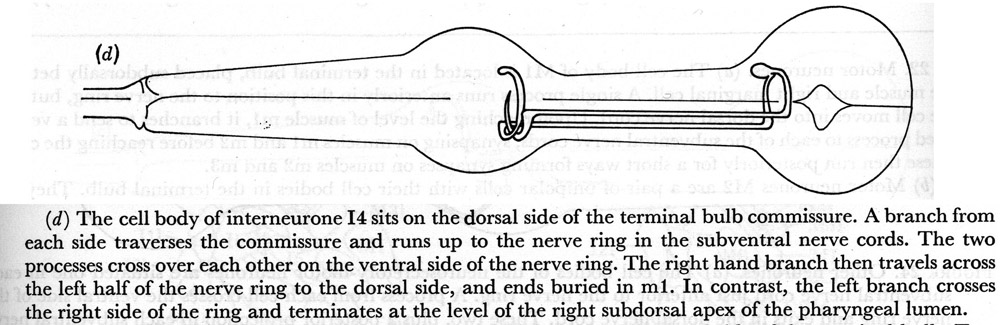
(d) The cell body of interneurone I4 sits on the dorsal side of the terminal bulb commissure. A branch from each side traverses the commissure and runs up to the nerve ring in the subventral nerve cords. The two processes cross over each other on the ventral side of the nerve ring. The right hand branch then travels across the left half of the nerve ring to the dorsal side, and ends buried in m1. In contrast, the left branch crosses the right side of the ring and terminates at the level of the right subdorsal apex of the pharyngeal lumen.
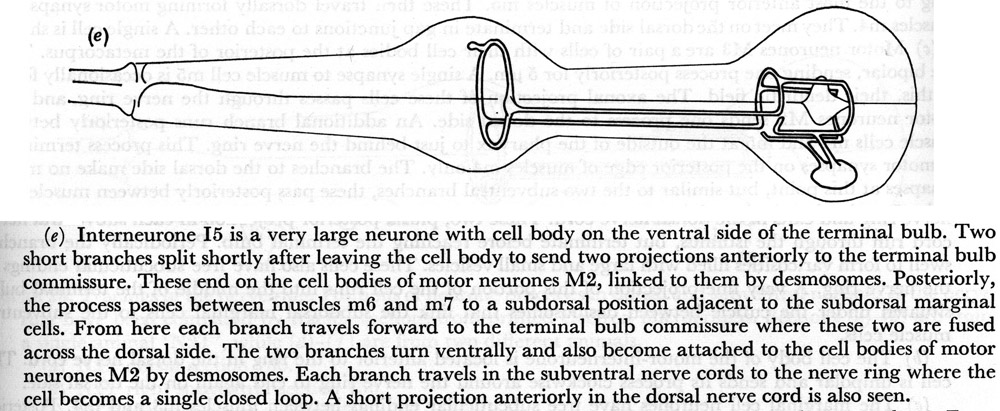
(e) Interneurone I5 is a very large neurone with cell body on the ventral side of the terminal bulb. Two short branches split shortly after leaving the cell body to send two projections anteriorly to the terminal bulb commissure. These end on the cell bodies of motor neurones M2, linked to them by desmosomes. Posteriorly, the process passes between muscles m6 and m7 to a subdorsal position adjacent to the subdorsal marginal cells. From here each branch travels forward to the terminal bulb commissure where these two are fused across the dorsal side. The two branches turn ventrally and also become attached to the cell bodies of motor neurones M2 by desmosomes. Each branch travels in the subventral nerve cords to the nerve ring where the cell becomes a single closed loop. A short projection anteriorly in the dorsal nerve cord is also seen.
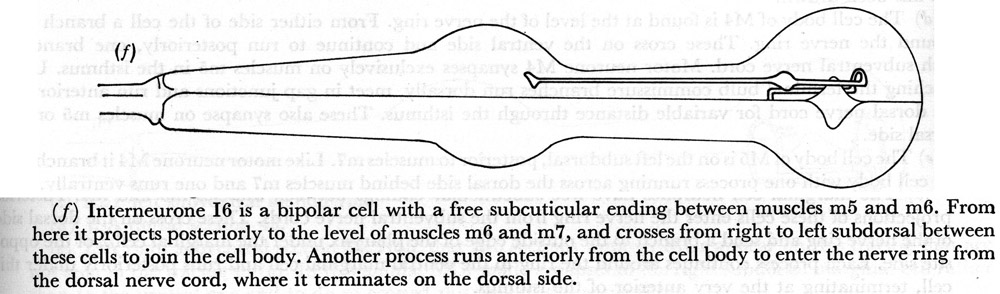
(f) Interneurone I6 is a bipolar cell with a free subcuticular ending between muscles m5 and m6. From here it projects posteriorly to the level of muscles m6 and m7, and crosses from right to left subdorsal between these cells to join the cell body. Another process runs anteriorly from the cell body to enter the nerve ring from the dorsal nerve cord, where it terminates on the dorsal side.
WA editors' note: The actual locations of cell bodies of neurons in the metacorpus are more posterior to those shown here (L. Avery pers. comm.)
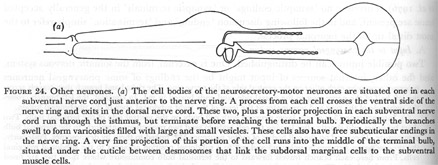
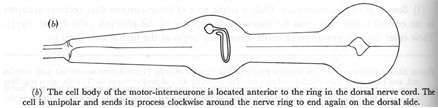
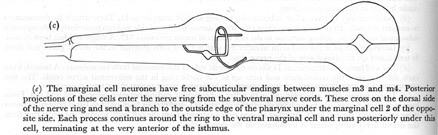 Figure 24. Other neurones.
Figure 24. Other neurones.
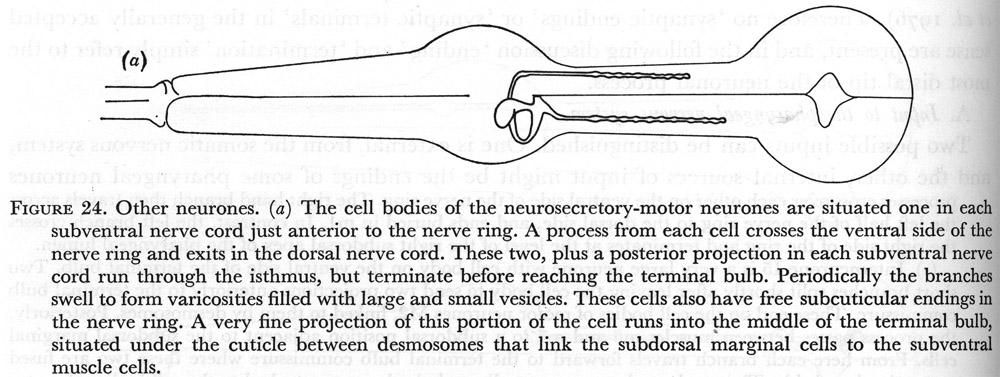
(a) The cell bodies of the neurosecretory-motor neurones are situated one in each subventral nerve cord just anterior to the nerve ring. A process from each cell crosses the ventral side of the nerve ring and exits in the dorsal nerve cord. These two, plus a posterior projection in each subventral nerve cord run through the isthmus, but terminate before reaching the terminal bulb. Periodically the branches swell to form varicosities filled with large and small vesicles. These cells also have free subcuticular endings in the nerve ring. A very fine projection of this portion of the cell runs into the middle of the terminal bulb, situated under the cuticle between desmosomes that link the subdorsal marginal cells to the subventral muscle cells.
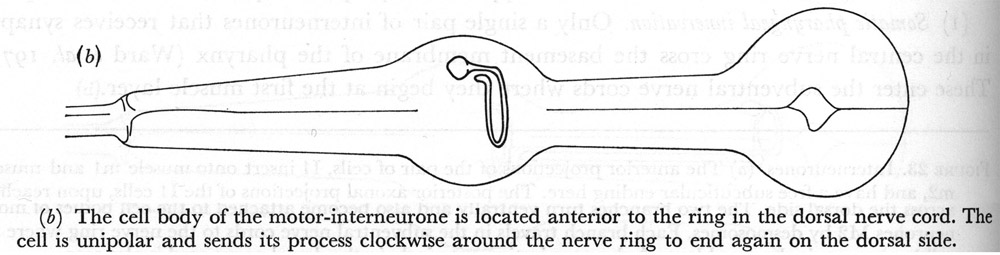
(b) The cell body of the motor-interneurone is located anterior to the ring in the dorsal nerve cord. The cell is unipolar and sends its process clockwise around the nerve ring to end again on the dorsal side.
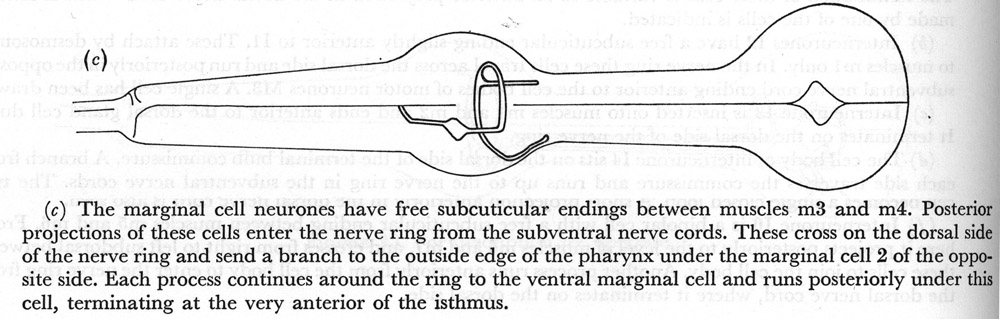
(c) The marginal cell neurones have free subcuticular endings between muscles m3 and m4. Posterior projections of these cells enter the nerve ring from the subventral nerve cords. These cross on the dorsal side of the nerve ring and send a branch to the outside edge of the pharynx under the marginal cell 2 of the opposite side. Each process continues around the ring to the ventral marginal cell and runs posteriorly under this cell, terminating at the very anterior of the isthmus.
WA editors' note: The actual locations of cell bodies of neurons in the metacorpus are more posterior to those shown here (L. Avery pers. comm.)
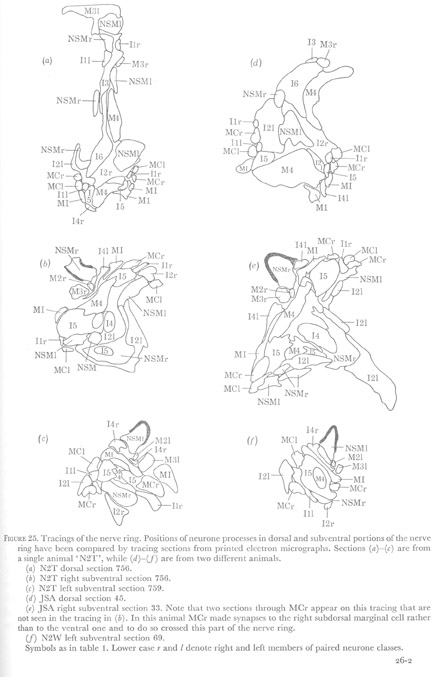 Figure 25. Tracings of the nerve ring. Positions of neurone processes in dorsal and subventral portions of the nerve ring have been compared by tracing sections from printed electron micrographs. Sections (a)-(c) are from a single animal 'N2T', while (d)-(f) are from two different animals.
Figure 25. Tracings of the nerve ring. Positions of neurone processes in dorsal and subventral portions of the nerve ring have been compared by tracing sections from printed electron micrographs. Sections (a)-(c) are from a single animal 'N2T', while (d)-(f) are from two different animals.
(a) N2T dorsal section 756.
(b) N2T right subventral section 759 (correction based on pers. comm with L. Avery).
(c) N2T left subventral section 759.
(d) JSA dorsal section 45.
(e) JSA right subventral section 33. Note that two sections through MCr appear on this tracing that are not seen in the tracing in (b). In this animal MCr made synapses to the right subdorsal marginal cell rather than to the ventral one and to do so crossed this part of the nerve ring.
(f) N2W left subventral section 69.
Symbols as in table 1. Lower case r and l denote right and left members of paired neurone classes.
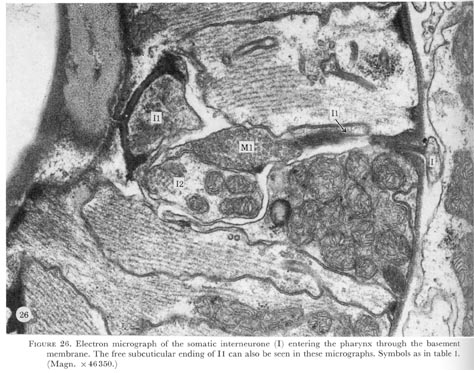 Figure 26. PLATE 8- Electron micrograph of the somatic interneurone (I) entering the pharynx through the basement membrane. The free subcuticular ending of I1 can also be seen in these micrographs. Symbols as in table 1. (Magn. X 46350.)
Figure 26. PLATE 8- Electron micrograph of the somatic interneurone (I) entering the pharynx through the basement membrane. The free subcuticular ending of I1 can also be seen in these micrographs. Symbols as in table 1. (Magn. X 46350.)
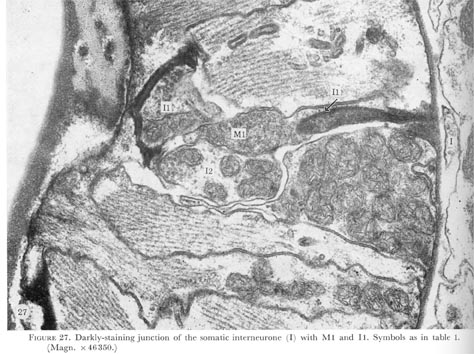 Figure 27. PLATE 8 - Darkly-staining junction of the somatic interneurone (I) with M1 and I1. Symbols as in table 1. (Magn. x 46350.)
Figure 27. PLATE 8 - Darkly-staining junction of the somatic interneurone (I) with M1 and I1. Symbols as in table 1. (Magn. x 46350.)
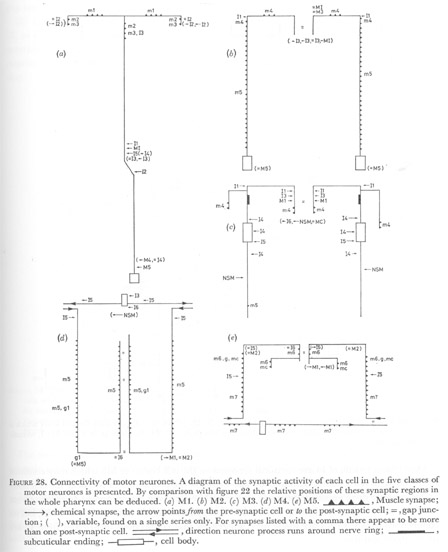 Figure 28. Connectivity of motor neurones. A diagram of the synaptic activity of each cell in the five classes of motor neurones is presented. By comparison with figure 22 the relative positions of these synaptic regions in the whole pharynx can be deduced. (a) M1. (b) M2. (c) M3. (d) M4. (e) M5.
Figure 28. Connectivity of motor neurones. A diagram of the synaptic activity of each cell in the five classes of motor neurones is presented. By comparison with figure 22 the relative positions of these synaptic regions in the whole pharynx can be deduced. (a) M1. (b) M2. (c) M3. (d) M4. (e) M5.  , Muscle synapse; , Muscle synapse;  , chemical synapse, the arrow points from the pre-synaptic cell or to the post-synaptic cell; = , gap junction; ( ), variable, found on a single series only. For synapses listed with a comma there appear to be more than one post-synaptic cell. , chemical synapse, the arrow points from the pre-synaptic cell or to the post-synaptic cell; = , gap junction; ( ), variable, found on a single series only. For synapses listed with a comma there appear to be more than one post-synaptic cell.  , direction neurone process runs around nerve ring; , direction neurone process runs around nerve ring;  , subcuticular ending; , subcuticular ending;  , cell body. , cell body.
(WA editors' note: Variable synapses between I2 and M1 neurons shown here are not included in the I2 neuron connectivity diagram below.)
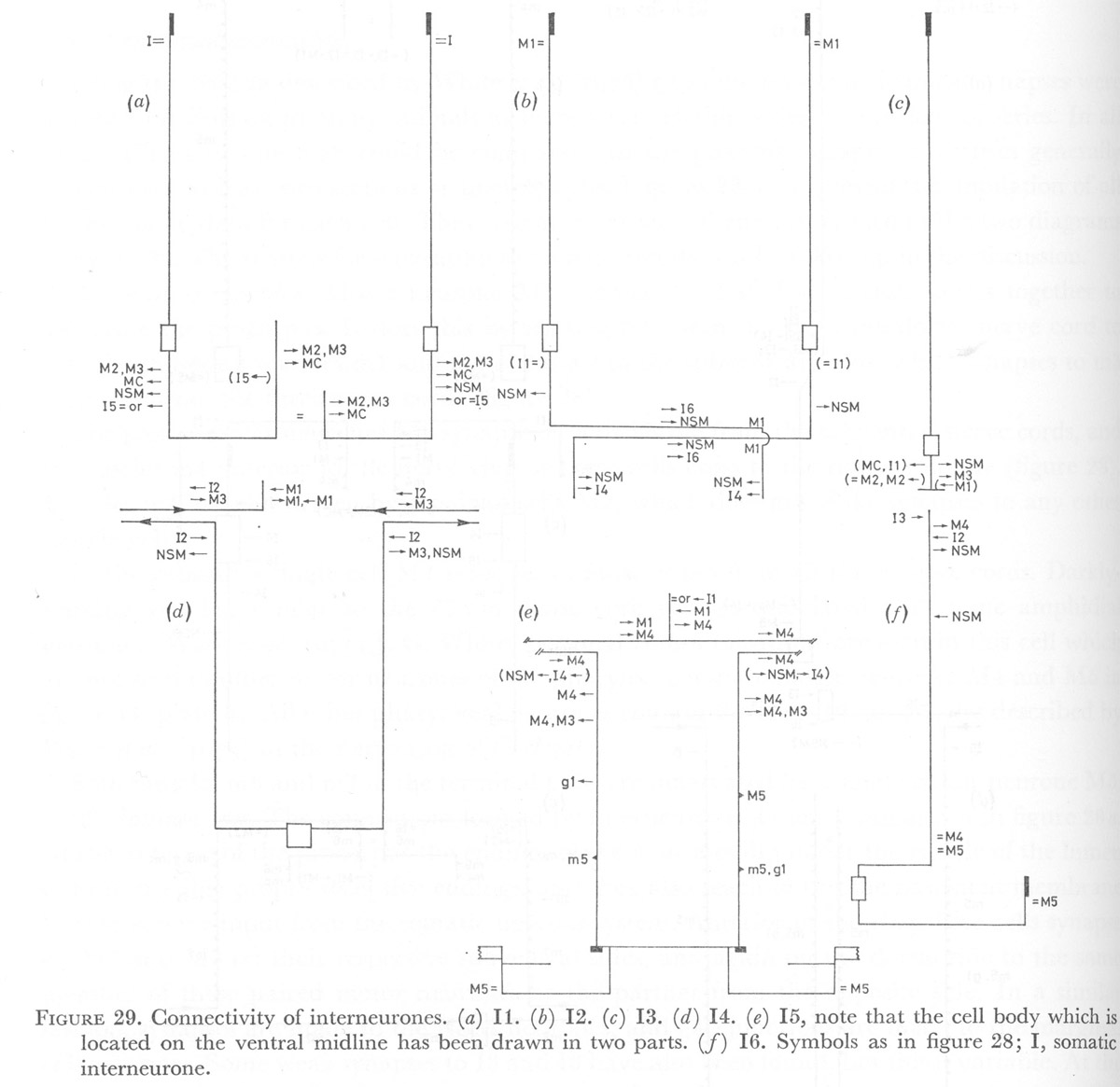
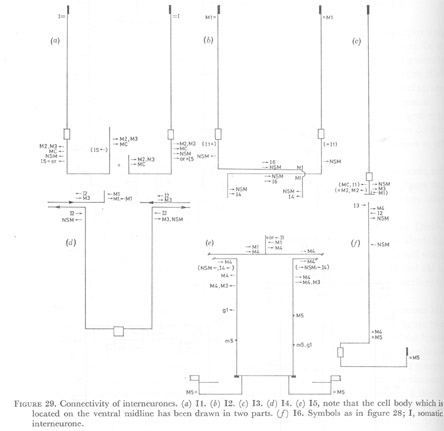 Figure 29. Connectivity of interneurones. (a) I1. (b) I2. (c) I3. (d) I4. (e) I5, note that the cell body which is located on the ventral midline has been drawn in two parts. (f) I6. Symbols as in figure 28; I, somatic interneurone.
Figure 29. Connectivity of interneurones. (a) I1. (b) I2. (c) I3. (d) I4. (e) I5, note that the cell body which is located on the ventral midline has been drawn in two parts. (f) I6. Symbols as in figure 28; I, somatic interneurone.
(WA editors' note: Variable synapses between I2 and M1 neurons shown in M1 diagram are not included in the I2 neuron diagram here.)
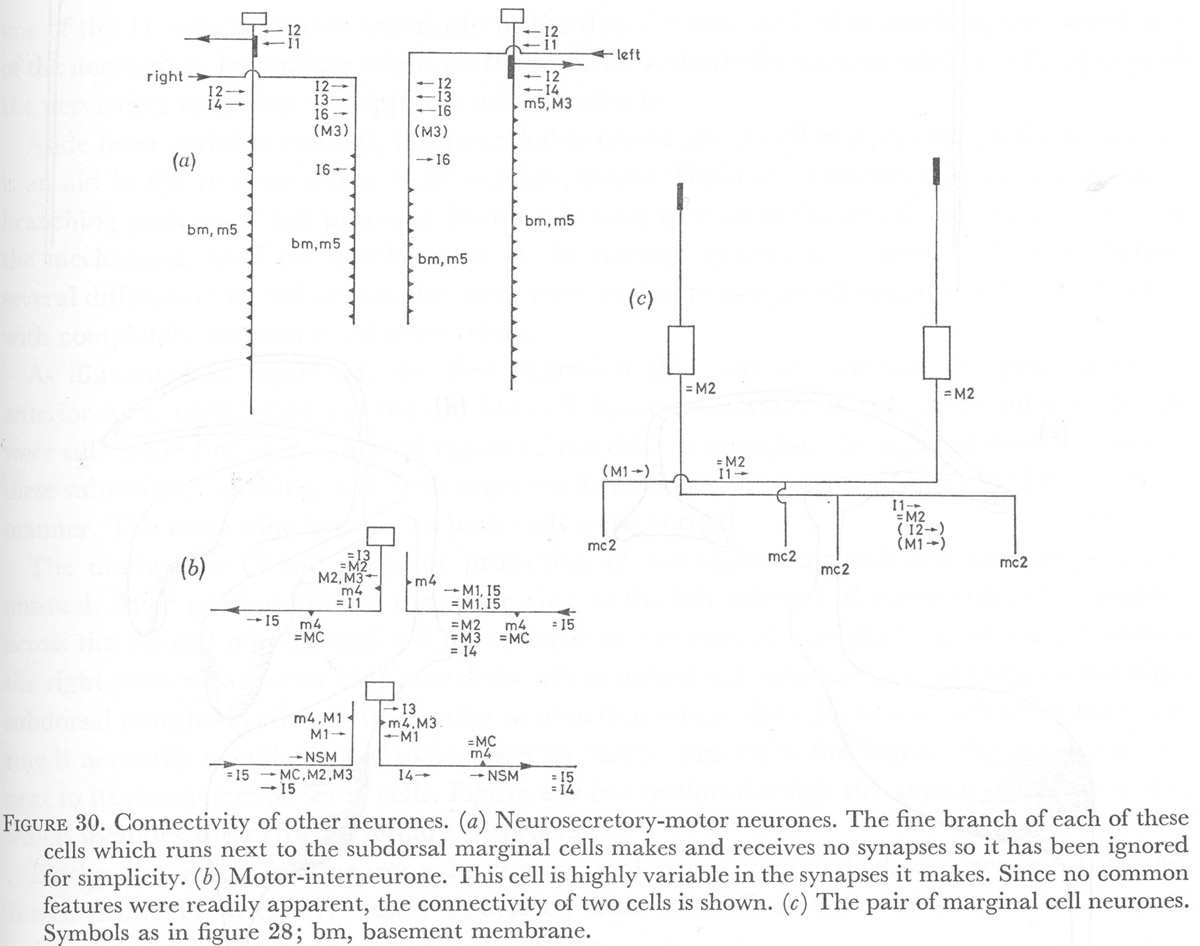
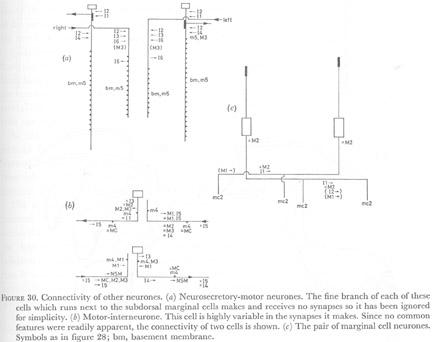 Figure 30. Connectivity of other neurones. (a) Neurosecretory-motor neurones. The fine branch of each of these cells which runs next to the subdorsal marginal cells makes and receives no synapses so it has been ignored for simplicity. (b) Motor-interneurone. This cell is highly variable in the synapses it makes. Since no common features were readily apparent, the connectivity of two cells is shown. (c) The pair of marginal cell neurones. Symbols as in figure 28; bm, basement membrane.
Figure 30. Connectivity of other neurones. (a) Neurosecretory-motor neurones. The fine branch of each of these cells which runs next to the subdorsal marginal cells makes and receives no synapses so it has been ignored for simplicity. (b) Motor-interneurone. This cell is highly variable in the synapses it makes. Since no common features were readily apparent, the connectivity of two cells is shown. (c) The pair of marginal cell neurones. Symbols as in figure 28; bm, basement membrane.
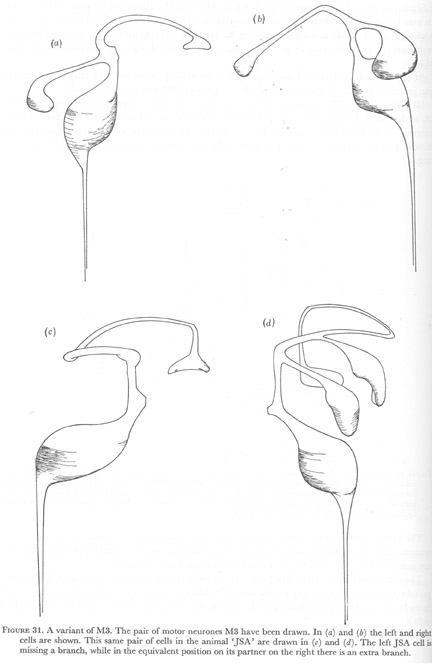 Figure 31. A variant of M3. The pair of motor neurones M3 have been drawn. In (a) and (b) the left and right cells are shown. This same pair of cells in the animal 'JSA' are drawn in (c) and (d). The left JSA cell is missing a branch, while in the equivalent position on its partner on the right there is an extra branch.
Figure 31. A variant of M3. The pair of motor neurones M3 have been drawn. In (a) and (b) the left and right cells are shown. This same pair of cells in the animal 'JSA' are drawn in (c) and (d). The left JSA cell is missing a branch, while in the equivalent position on its partner on the right there is an extra branch.
Discussion
(a) Comparitive anatomy
(b) Comparison of pharyngeal and body neuromuscular systems
(c) Patterns of innervation
(d) Growth of neurones
(e) Multi-functional cells
(f) Behaviour
(i) Pumping
(ii) Control of pumping
(a) Comparative anatomy
The nematode pharynx has been used widely as the basis for the taxonomy of these animals, because nuclei, organized in well-defined and constant arrays were easily visualized by light microscopists. If the distribution of nuclei in C. elegans displayed in figure 14 is compared with the charts for Rhabditis worked out by Chitwood & Chitwood (1938), then one sees the basic plan is the same. However, in the present work several of the nuclei in the corpus nerve cords were found to be epithelial or muscle cell bodies, rather than neurones, as they had been assigned previously.
The electron microscope has shed some further light on the question of the syncytial nature of the organ. As reviewed by Bird (1971), early workers had failed to find cell membranes in the pharynx, and more recently Reger (1966) described Ascaris as having a syncytial myoepithelium in the pharynx. This complete reconstruction of the pharynx and other electron microscopic studies (Lee 1968; Yuen 1968b) clearly demonstrate that cell membranes can be observed in many species. However, it is worth noting that muscle cells in C. elegans are binucleate within sectors, dividing up generally into layers of three cells. In some places where cells are free to cross such boundaries as the marginal cells, the cells do fuse. This is seen on the ventral side of muscles m5 and for the dorsal and right g1 gland cells. Why in this last instance only the right side should become fused is unclear, but this was observed in the three animals examined. This pattern of fusion contrasts with observations on other species for which Yuen (1968a) found the subventral glands of Ditylenchus were fused in the anterior half of the terminal bulb, while much earlier Looss had described the fusion of all three gland cells of Ancyclostoma (Bird 1971). One is led to the conclusion that there is a tendency towards a syncytium in the pharynx of nematodes. In contrast, in the body of C. elegans only the hypodermal cells appear to behave similarly (White et al. 1976).
(b) Comparison of pharyngeal and body neuromuscular systems
Structurally body wall and pharyngeal muscle are different. Body muscles run longitudinally and are 'obliquely striated' (Rosenbluth 1965), while each pharyngeal muscle sector is composed of a single sarcomere running between basement membrane and cuticle. Biochemical differences in myosin heavy chains, as well as the isolation of mutants affecting only the body muscle myosin further distinguishes these two muscle classes (Epstein, Waterston & Brenner 1974).
Neuromuscular relations in the pharynx also differ from the somatic system. As described in the accompanying paper (White et al. 1976), the body muscles have muscle arms that extend to the nerve cords to receive synapses. In the pharynx neurones run within muscle sectors and most synapses to muscle are made en passant. Motor neurones M3 are unusual in this regard, sending a branch to the most posterior projection of muscles m4 to make their synapses.
(c) Patterns of innervation
The pharynx is a three part structure. In contrast, the pharyngeal neurones are paired subventral cells or they are single dorsal cells, often with two equivalent branches. In the case of motor neurones, paired subventral cells (M2, M3) form muscle synapses from the subventral nerve cords and then run around to the dorsal muscle sector where both cells synapse onto the dorsal muscle cell. Single dorsal motor neurones, such as M4 and M5, have one branch in each subventral nerve cord synapsing onto muscles; both branches then travel to the dorsal side to synapse onto the dorsal muscle sector. Thus, by having the dorsal sector receive synapses from both paired subventral cells or both branches of dorsal cells, the nervous system can innervate the three muscle sectors of the pharynx. In a similar fashion, the marginal cell neurones each synapse onto one subdorsal marginal cell and both neurones synapse onto the ventral cell. Also, the paired subventral interneurones synapse onto target cells on the subventral side of the nerve ring and onto the same cell and/or its partner of paired target cells on the dorsal side. Thus, the neurone classes in the pharynx are bilaterally symmetric about the dorsoventral midline. For symmetry in the single dorsal cells with two branches a line must be drawn through the cell body, so that each branch of the cell plus one-half nucleus behaves similarly to one of the paired subventral neurones. Possibly, this might suggest that these single dorsal cells are tetraploid.
(d) Growth of neurones
Termination of neuronal growth in the pharynx may occur by fibres meeting and forming gap junctions. Thus, when pairs of cells or branches of a single cell end on the dorsal midline, they do so by meeting in gap junctions to each other. In the preceding paper, similar behaviour occurring between classes of ventral cord neurones is described (White et al. 1976).
Motor neurones in the pharynx specifically innervate particular classes of target cell. For example, the motor neurones M3 make motor synapses on the posterior projection of m4. Here the neurone lies between two muscle cells m4 and m5, but synaptic bars on the m5 side of the neurone are never seen. The one exceptional animal in which several changes were seen in the nervous system provides another example of the preference neurones have for certain target cell classes. When the marginal cell neurone failed to synapse on its normal target, the ventral marginal cell, it continued to grow around the nerve ring until it eventually synapsed on another marginal cell, rather than on muscles or neurones.
Paths of cells in the nerve ring might be laid down by other cells. This is suggested from the reconstruction of the one exceptional animal. When a pair of cells that lie one on top of the other in the nerve ring were both found to have altered morphology in the ring it indicated that one cell (I4) could take its growth cues from the other (MI) and follow along this fibre, possibly recognizing a membrane polarity which leads to its asymmetric growth. If a range of alterations in the pharyngeal nervous system could be collected by isolation of mutants then further mechanisms for specifying neuronal growth might be deduced.
(e) Multi-functional cells
Several neurone classes in the pharynx anatomically appear to have several functions, aside from their synaptic assignments as either motor or interneurones. Many have free subcuticular endings suggesting that they are also mechanoreceptors. Motor neurones M3 could therefore be sensory-motor neurones, similar to other neurones described in the somatic nervous system of C. elegans (Ward et al. 1975). The NSM cells have been called secretory because of the large vesicles seen in these cells and so could be not only sensory-motor neurones, but neurosecretory cells, as well. Cells with the ultrastructure of neurosecretory neurones were also observed by Lee (1968) in studies of the pharynx of Nippostrongylus brasiliensis. Possibly such cells in the pharynx of C. elegans, which, because they lie at the outside edge of the pharynx, might secrete a humoral factor into the pseudocoelom when their sensory endings detect food in the lumen.
In addition to multi-functional neurones, the muscles m6 appear both to be contractile cells, and to lay down the cuticle in the pharynx. Vesicles are seen at the grinder side of m6 and in some animals the vesicles were filled with material that looked similar to the cuticle. It appeared that cuticular matter was being exocytosed toward the cuticle of the grinder at the time of fixation.
Multi-functional cells, in particular sensory-motor neurones, have been suggested as intermediates in the evolution of nervous systems (Horridge 1968 ; Coggeshall 1971). Thus, the pharynx of C. elegans appears to be not only a simple neuromuscular system, but also a primitive one.
(f) Behaviour
By using Nomarski optics in the light microscope, the movements of individual pharyngeal muscle layers can be observed in animals feeding on bacteria. When observations were made in this way on feeding C. elegans, it was found that the description of feeding given by Doncaster (1962) for Rhabditis oxycerca and Pelodera lambdiensis also applied to C. elegans. Briefly, in each cycle of pumping the grinder is set in motion first, when the 'flaps' are pulled backwards by muscle contraction in the terminal bulb causing food in front of the grinder to be processed through. Then the corpus opens, as well as the anterior part of the isthmus, and food particles and liquid are sucked in. This region closes again before the contraction stroke of the terminal bulb has progressed to the point of causing ground food to pass in to the gut. After this, the grinder flaps return to the resting, T-shaped configuration and there is a slight pause before they start to move forward again, initiating a new cycle of the pump. Particulate food is trapped by the corpus, while excess water is expelled when the pharynx closes. Food plugs accumulate in the anterior half of the isthmus, and then portions are passed backwards through the isthmus to lie just in front of the grinder.
The nematode feeds almost constantly during its life cycle, except during the moult, or when a special, resistant form, the dauer larva, is induced upon starvation. At these times a mechanism for inhibiting pharyngeal pumping is required. Furthermore, during the end of the moult, the corpus alone must be inhibited while at least the back half of the pharynx is activated for ingestion of the shed cuticle.
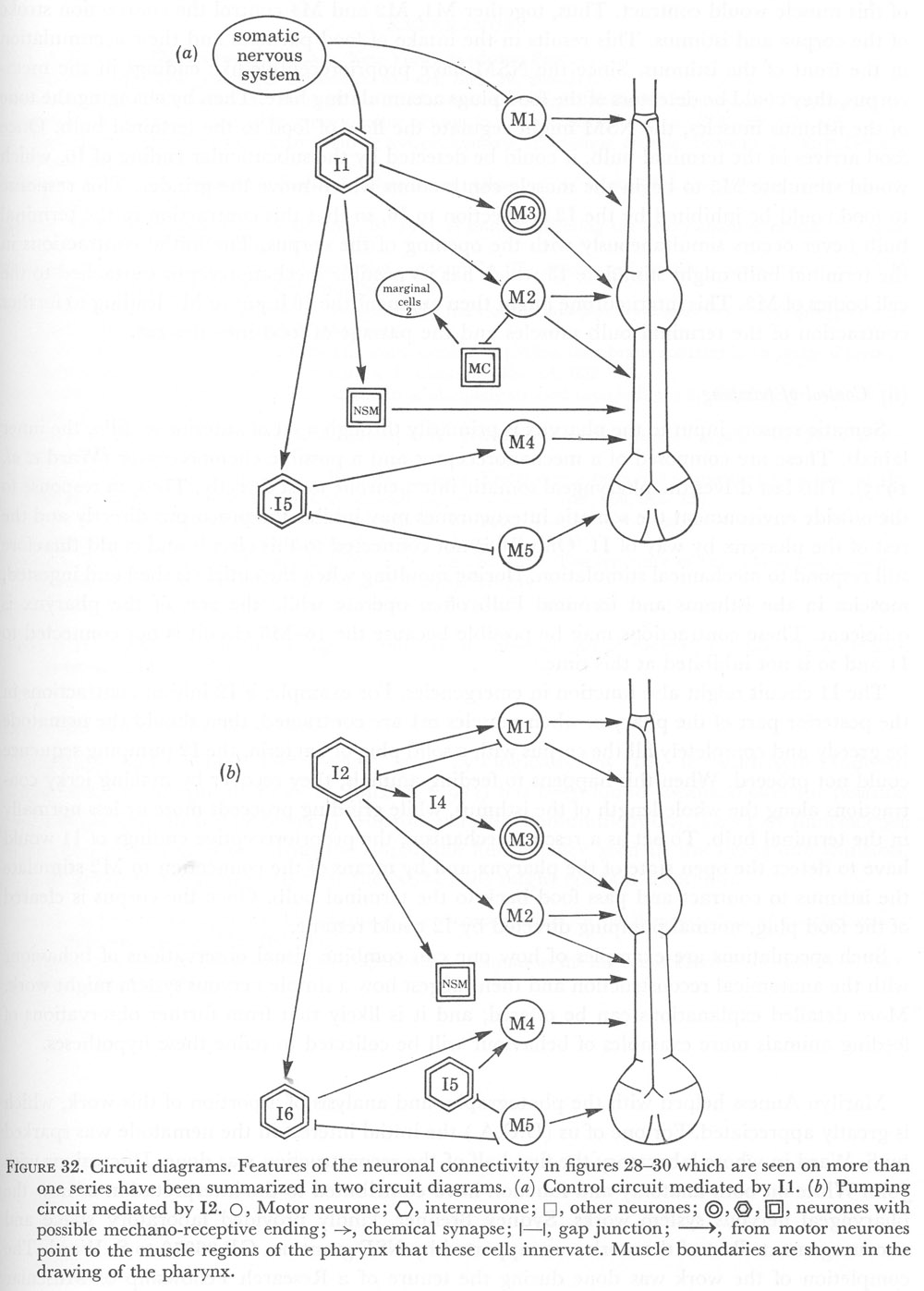
It seems likely that there are two different pathways for controlling pumping and its inhibition. The diagrams in figure 32 indicate a possible mechanism in which it is suggested that I1 mediate inhibition, since these cells receive input from the somatic nervous system. Neurones I2, which have connections to all motor neurones would then control pumping.
(i) Pumping
Interneurones I2 are not connected to the somatic nervous system, and therefore each cycle of the pump must be regenerated from within the pharynx itself. The I2 cells have proprioreceptive-like endings attached to muscle m1 and could excite M1 and initiate opening of the procorpus when it is closed. This would start a cycle of the pump. At the same time as the procorpus opens, the metacorpus and front of the isthmus would be opened by I2 exciting M2 to cause contraction of muscles m4 and m5.
However, motor neurones M2 make synapses along the length of the single isthmus muscle m5, suggesting that the whole of this muscle should contract in response to M2, but during normal pumping only the anterior part opens. An inhibitory motor neurone in the isthmus, acting antagonistically to M2 might modulate the contraction of m5. Motor neurone M4 appears anatomically ideally suited to this. Examination of the distribution of motor synapses from M4 reveals that at the anterior of the isthmus the branches of M4 are dendritic while motor synapses are concentrated posteriorly in the isthmus. Here also, all three muscle sectors are innervated by the two subventral and two dorsal branches making muscle synapses (figure 28). Thus, inhibition from M4 would be concentrated at the back of muscles m5, and if in this region inhibition is greater than the excitation from M2, then only the anterior portion of this muscle would contract. Thus, together M1, M2 and M4 control the contraction stroke of the corpus and isthmus. This results in the intake of food particles and their accumulation in the front of the isthmus. Since the NSM have proprioreceptive-like endings in the metacorpus, they could be detectors of the food plugs accumulating here. Then by changing the tone of the isthmus muscles, the NSM might regulate the flow of food to the terminal bulb. Once food arrives in the terminal bulb, it could be detected by the subcuticular ending of I6, which would stimulate M5 to begin the muscle contractions which move the grinder. This response to food could be inhibited by the I2 connection to I6, so that this contraction in the terminal bulb never occurs simultaneously with the opening of the corpus. The initial contractions in the terminal bulb might stimulate I5 which has its possible mechanoreceptors attached to the cell bodies of M2. This interneurone might then augment the I6 input to M5 leading to further contraction of the terminal bulb muscles and the passage of food into the gut.
(ii) Control of pumping
Somatic sensory input to the pharynx is primarily through a set of anterior sensilla, the inner labials. These are composed of a mechanoreceptor and a possible chemoreceptor (Ward et al. 1975). This last drives the pharyngeal somatic interneurone most directly. Thus, in response to the outside environment the somatic interneurones may inhibit the procorpus directly and the rest of the pharynx by way of I1. Only I6 is not connected to this circuit and could therefore still respond to mechanical stimulation. During moulting when the cuticle is shed and ingested, muscles in the isthmus and terminal bulb often operate while the rest of the pharynx is quiescent. These contractions may be possible because the I6-M5 circuit is not connected to I1 and so is not inhibited at this time.
The I1 circuit might also function in emergencies. For example, if I2 inhibit contractions in the posterior part of the pharynx when muscles m1 are contracted, then should the nematode be greedy and completely fill the corpus with a solid plug of bacteria, the I2 pumping sequence could not proceed. When this happens to feeding animals, they recover by making jerky contractions along the whole length of the isthmus, while grinding proceeds more or less normally in the terminal bulb. To act as a rescue mechanism, the proprioreceptive endings of I1 would have to detect the open state of the pharynx and by means of the connection to M2 stimulate the isthmus to contract and pass food back to the terminal bulb. Once the corpus is cleared of the food plug, normal pumping directed by I2 could resume.
Such speculations are examples of how one can combine visual observations of behaviour with the anatomical reconstruction and then suggest how a simple nervous system might work. More detailed explanations can be offered, and it is likely that from further observations of feeding animals more examples of behaviour will be collected to refine these hypotheses.
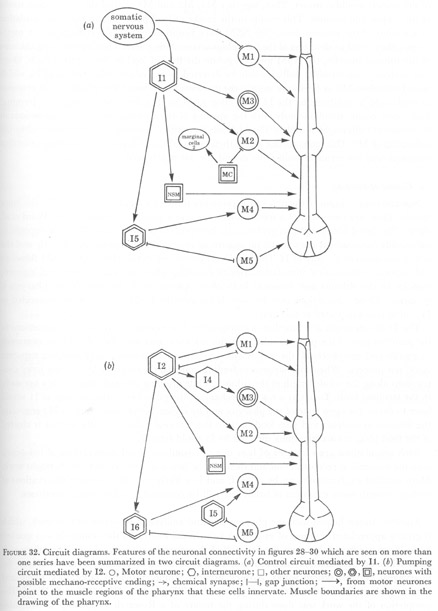 Figure 32. Circuit diagrams. Features of the neuronal connectivity in figures 28-30 which are seen on more than one series have been summarized in two circuit diagrams. (a) Control circuit mediated by I1. (b) Pumping circuit mediated by I2.
Figure 32. Circuit diagrams. Features of the neuronal connectivity in figures 28-30 which are seen on more than one series have been summarized in two circuit diagrams. (a) Control circuit mediated by I1. (b) Pumping circuit mediated by I2.  , Motor neurone; , Motor neurone;  , interneurone; , interneurone;  , other neurones; , other neurones;  , neurones with possible mechano-receptive ending; , neurones with possible mechano-receptive ending;  , chemical synapse; , chemical synapse;  , gap junction; , gap junction;  , from motor neurones point to the muscle regions of the pharynx that these cells innervate. Muscle boundaries are shown in the drawing of the pharynx. , from motor neurones point to the muscle regions of the pharynx that these cells innervate. Muscle boundaries are shown in the drawing of the pharynx.
Acknowledgments
Marilyn Anness helped with the photography and analysis of a portion of this work, which is greatly appreciated. For one of us (D.G.A.) the initial interest in the nematode was sparked by S. Ward in whose laboratory the first half of the reconstruction was done. Discussions with John White on both anatomy and function have contributed to the interpretation of how the pharyngeal nervous system works. Sydney Brenner, kindly provided laboratory space and encouragement. Part of the work was supported by NSF grant no. GB 36970 to S. Ward. The completion of the work was done during the tenure of a Research Fellowship of Muscular Dystrophy Associations of America to D.G.A.
References
Bird, A. F. 1971 The structure of nematodes. New York: Academic Press.
Brenner, S. 1973 The genetics of behaviour. Br. med. Bull. 29, 269-271.
Chitwood, B. G. & Chitwood, M. B. 1938 An introduction to nematology. Baltimore: Monumental Printing Co.
Coggeshall, R. E. 1971 A possible sensory-motor neuron in Aplysia California. Tissue & Cell 3, 637-648.
de Coninck, L. 1965 Généralités. In Traité de zoologie, anatomie, systématique, biologie, Tome IV (ed. P.-P. Grassé). Paris: Masson et Cie.
Doncaster C. C. 1962 Nematode feeding mechanisms. I. Observations on Rhabditisand Pelodera. Nematologica 8, 313-320.
Epstein, H. F., Waterston, R. F. & Brenner, S. 1974 A mutant affecting the heavy chain of myosin in Caenorhabditis elegans. J. molec. Biol. 90, 291-300.
Horridge, G. A. 1968 The origins of the nervous system. In The structure and function of nervous tissue (ed. G. H. Bourne), vol. I. New York : Academic Press.
Lee, D. L. 1968 The ultrastructure of the alimentary tract of the skin-penetrating larva of Nippostrongylus brasiliensis (Nematoda). J. Zool., Lond. 154, 9-18.
Reger, J. 1966 The fine structure of fibrillar components and plasma membrane contacts in esophageal myoepithelium of Ascaris lumbricoides (var. suum). J. Ultrastruc. Res. 14, 602-617.
Rosenbluth, J. 1965 Ultrastructural organisation of obliquely striated muscle fibres in Ascaris lumbricoides. J. Cell Biol. 25, 495-515.
Sulston, J .E. 1976 Post embryonic development in the ventral cord of Caenorhabditis elegans. Phil. Trans. R. Soc. Lond. B 275, 287-297.
Ward, S., Thomson, N., White, J .G. & Brenner, S. 1975 Electron microscopical reconstruction of the anterior sensory anatomy of the nematodeCaenarhabditis elegans. J. camp. Neur. 160, 313-338.
Ware, R. W., Clark, D., Crossland, K. & Russell, R. L. 1975 The nerve ring of the nematode Caenorhabditis elegans: sensory input and motor output. J. camp. Neur. 162, 71-110.
White, J .G., Southgate, E., Thomson, J. N. & Brenner, S. 1976 The structure of the ventral nerve cord of Caenorhabditis elegans. Phil. Trans. R. Soc. Lond. B 275, 327-348.
Yuen, P. H. 1968a Electron microscopical studies on Ditylenchus dipsaci. II. Oesophagus. Nematalogica 14, 385-394.
Yuen, P. H. 1968b Electron microscopical studies on the anterior end of Panagrellus silusiae(Rhabditidae). Nematalogica 14, 554-564.
Note added in proof, 20 April 1976
Recently, pale yellow fluorescence seen cytologically by the formaldehyde induced fluorescence technique of Fuxe & Jonsson (1973) has been found localized over the position of the NSM cells in the pharynx (J. Sulston, personal communication). This pale fluorescence which can be distinguished from the dopamine fluorescence seen in the nematode (Sulston, Dew & Brenner 1975) suggests that the NSM cells might contain serotonin.
Fuxe, K. & Jonsson, G. 1973 The histochemical fluorescence method for the demonstration of catecholamines. J. Histochem. Cytochem. 21,293-311.
Sulston, J., Dew, M. & Brenner, S. 1975 Dopaminergic neurons in the nematode Caenorhabditis elegans. J. comp. Neur. 163, 215-226.
|