|
EPITHELIAL SYSTEM OF THE MALE - SEAM AND TAIL HYPODERMIS
 Click pictures for new window with figure and legend Click pictures for new window with figure and legend
1 The Seam
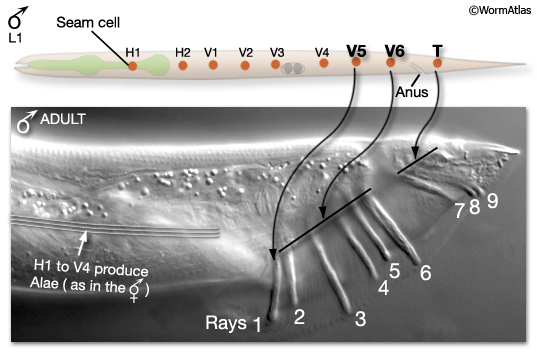
In the male, the three most posterior L1 lateral seam cells, V5 (L/R), V6 (L/R) and T (L/R), produce the sensory rays, instead of alae-secreting seam cells (MaleHypFIG 1). Rays are required for various steps in male mating behavior (to view behavior see Male Introduction).
MaleHypFIG 1: L1 seam cells V5, V6 and T produce rays in the male.
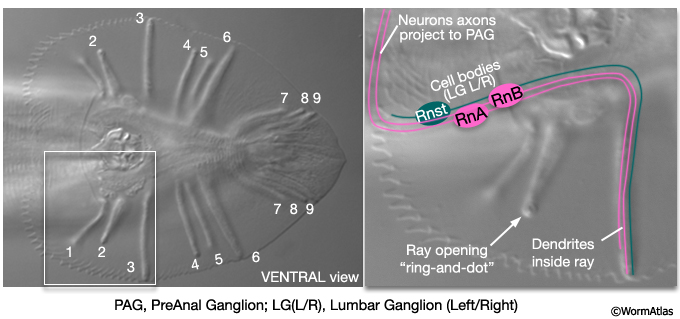
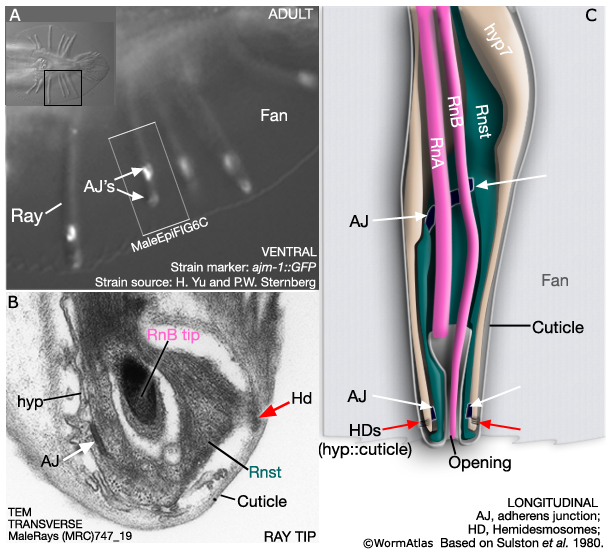
The adult male tail bears a total of nine bilateral pairs of rays (numbered, n, 1 to 9 from anterior to posterior, left to right) which are embedded in a cuticular fan (MaleHypFIG 2). Each ray consists of the dendritic endings of two sensory neurons (an RnA and an RnB neuron), surrounded by an epithelial tube formed by a ray structural cell (Rnst). All ray cell processes are ensheathed in an extension of hyp7 and cuticle (MaleHypFIG 2; for details see Male Neuronal Support Cells - Rays). Except for ray 6, the tips of the rays open to the environment. At light microscope resolution, ray openings can be seen as a "ring-and-dot", formed by the hyp and Rnst cell encircling the RnB neuron tip. Ray development spans L2 to late L4 and can be divided into two non-overlapping phases (1) ray cell generation and (2) ray morphogenesis.
MaleHypFIG 2: Ray structure.
|
1.1 Ray Cell Generation
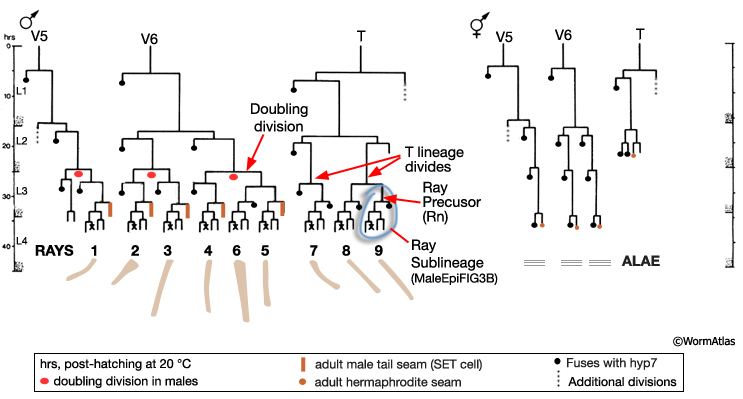
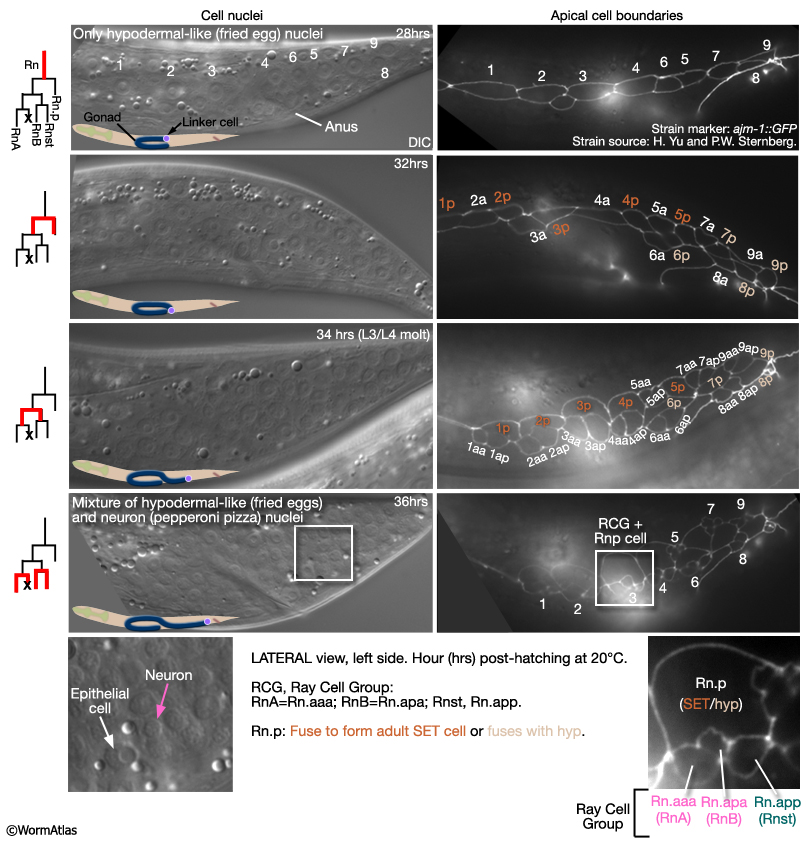
The first signs of sexual dimorphism in the seam are apparent at L2 where male V5 and V6 lineages execute a doubling division and the T lineage divides (MaleHypFIG 3A). The daughters of the V5 and V6 doubling division, and T, divide to produce 9 ray precursor (Rn) cells in each seam by L3 (MaleHypFIG 4, top row).
MaleHypFIG 3A: Male versus hermaphrodite seam lineages.
MaleHypFIG 3B: The ray sublineage.
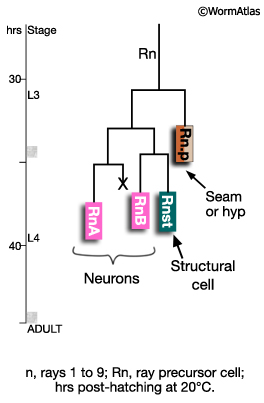
Between mid-L3 and mid-L4, each Rn cell divides by a stereotyped lineage pattern (referred to as the ray sublineage) producing the constituent cells of a single ray: an RnA, RnB and Rnst cell (a Ray Cell Group, RCG) (MaleHypFIG 3B and MaleHypFIG 4). Thus, the cells of a single ray derive from a common precursor. Execution of the sublineage is driven by members of the conserved basic-helix-loop-helix proneural gene family (LIN-32 and HLH-2; Zhao and Emmons, 1995; Portman and Emmons, 2000) and a DM domain gene (MAB-3; Yi et al. 2000; Ross et al, 2005). In addition to a RCG, each Rn produces 1 hypodermal cell (Rn.p) that will either fuse with Rn.p cells from other rays forming the male posterior seam (SET) cell (R1.p-R5.p) or will fuse with hyp 7 (R6.p-R9.p; Sulston et al., 1980).
MaleHypFIG 4: Ray cell generation.
1.2 Ray Morphogenesis
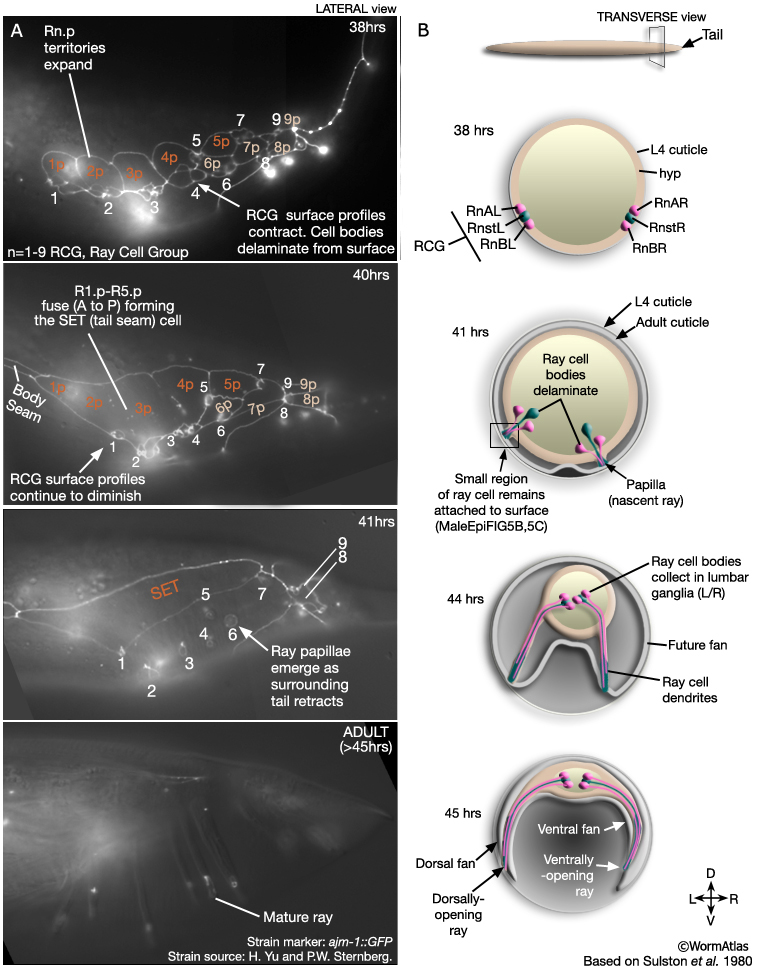
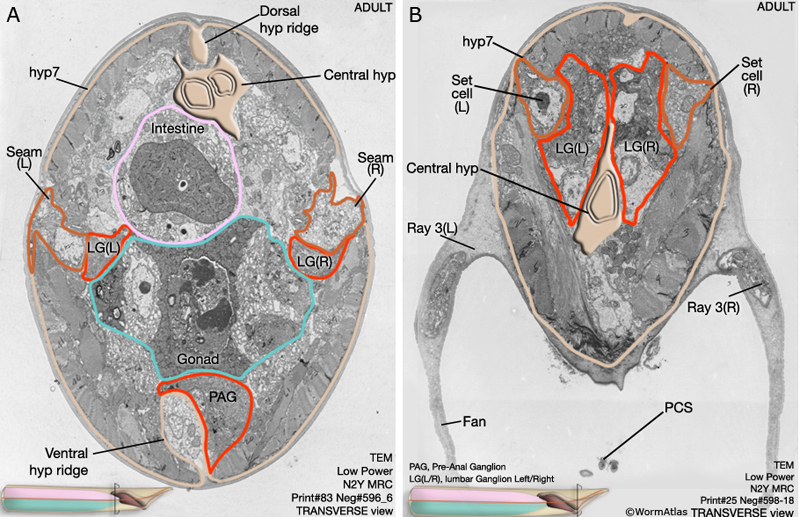
After completion of the ray sublineages the hypodermis, ray cell bodies and other cells of the posterior migrate anteriorly as part of the process of reshaping the male tail. Ray cell bodies detach from the body wall and migrate anteriorly and medially to the lumbar ganglia (LG L/ R) (MaleHypFIG 5B and MaleHypFIG 8). A small part of each ray cell, however, remains attached to the body wall and the general forward movement of hyp and ray cell bodies spins out the rays (MaleHypFIG 6B&C).
MaleHypFIG 5: Lateral and transverse views of ray morphogenesis.
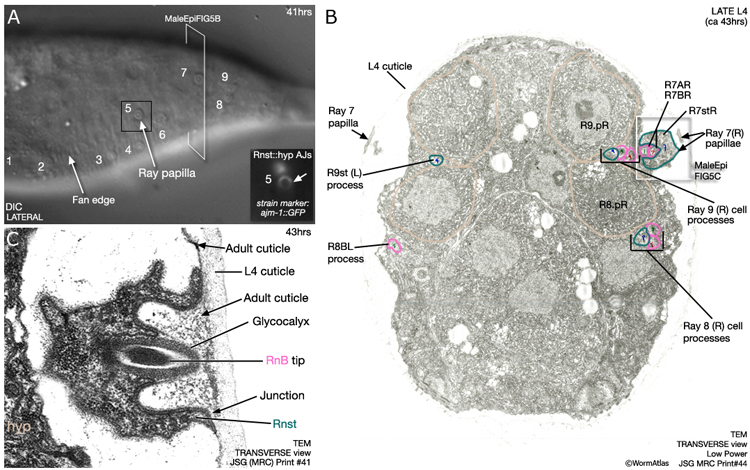
Nascent rays initially appear as papillae on the surface of the tail (MaleHypFIG 6) and subsequently lengthen as surrounding tissues continue to move forward. The fan forms from cuticle that separates from the body wall at the lateral sides of the animal (MaleHypFIG 5B). The gap formed is bridged by the developing rays. The inner layer of the adult cuticle is laid down and the lateral portions of cuticle collapse around the rays forming the fan. The ray tips characteristically open on either the dorsal (rays 1, 5 and 7) or ventral (rays 2, 4 and 8) fan surface or at the fan margin (rays 3 and 9) (Sulston et al., 1980) and this is established when the ray papillae initially attach to the cuticle (as shown in MaleHypFIG 6B&C).
MaleHypFIG 6: Ray papilla.
As mentioned above, a small part of each ray cell is connected to the body wall and these fixed points of attachment, coupled with retraction of surrounding tissues, drive ray formation. Ultrastructurally the attachments consist of adherens junctions (AJs) interlinking hyp, Rnst and ray neurons as well as hemidesmosomes (HDs), that link hyp to cuticle (MaleHypFIG 7A-C).
MaleHypFIG 7: Ray ultrastructure.
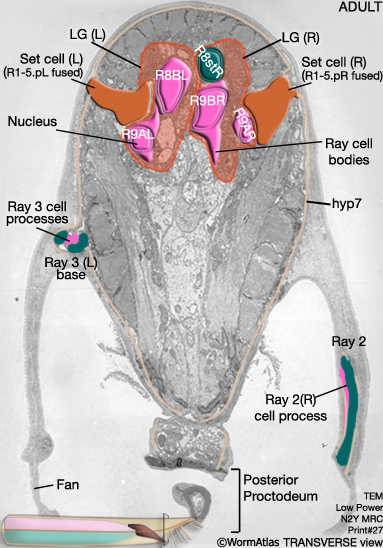
MaleHypFIG 8: EM cross-section of the adult male tail.
During the course of ray cell generation and morphogenesis some cell-cell contacts are maintained while others change (Baird et al., 1991). For example, the constituent cells of the RCG remain in tight association throughout ray development and do not intermingle with cells of other rays (MaleHypFIG 5A). This is reminiscent of the phenomenon of lineage restriction, observed in the Drosophila imaginal disc compartments or in the rhombomeres of vertebrate hindbrain. By contrast, RCGs must separate from the SET in order for future rays adopt their adult axial positions. Cell affinities and migrations are regulated by conserved cell attraction and repulsion factors such as ephrins, semaphorins and plexins (Roy et al, 2000; Ginzburg et al, 200; Hahn and Emmons, 2003; Dalpé et al, 2004). Mutations in genes encoding such factors cause abnormal ray positioning or fusion of rays (for EM analysis of fused ray ultrastructure see Chow et al., 1995).
2 The Tail Tip hyp
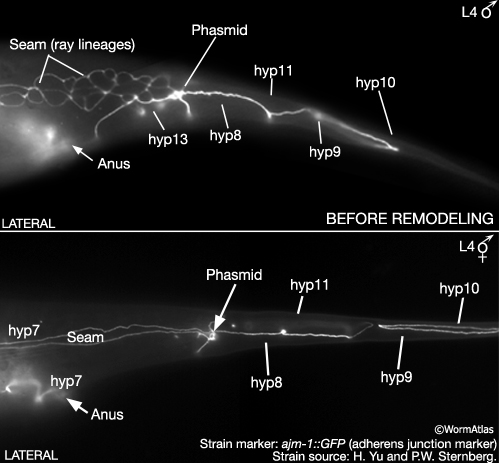
During late L4 the male tail tip changes from a tapering cone shape (leptoderan) to a blunt dome (peloderan) (MaleEpiFIG 1; for animation visit Fitch lab website). Tail tip remodeling is the first step in establishing the adult male posterior form. Prior to tip morphogenesis, male and hermaphrodite tail tips differ only slightly in their cell contacts and composition (MaleHypFIG 9). In both sexes the tip contains the 4 hyp cells hyp8-11. In males, however, the sisters of T form a binucleate cell called hyp13, instead of fusing with hyp7 as in the hermaphrodites (Sulston et al., 1983). Cell contacts are largely the same except that in males hyp11 borders with male-specific ray lineage cells and hyp8 with hyp13. The distribution and frequency of adherens junctions (AJs) and gap junctions (GPs) between tail tip cells does not differ significantly between the sexes at this stage (Nguyen et al., 1999).
MaleHypFIG 9: Male and hermaphrodite tails at L4.
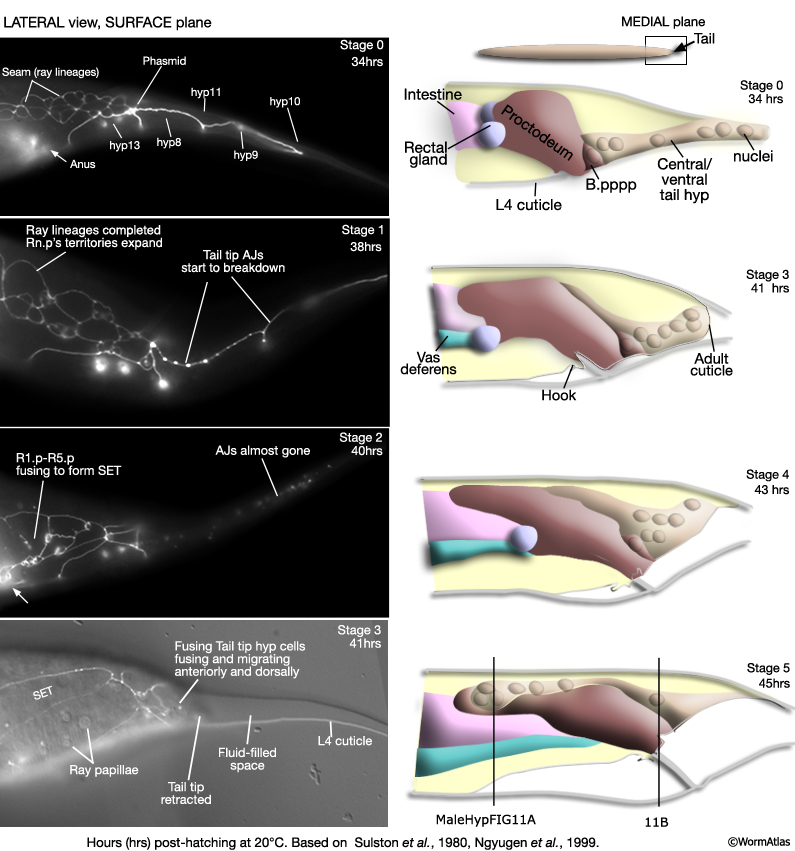
Tail tip remodeling occurs at the same time as ray morphogenesis (beginning mid L4, ca 35 hrs; MaleHypFIG 10). The process can be divided into approximately 5 stages (Sulston et al., 1980; Nguyen et al., 1999). Cell borders break down (visualized by the disappearance of adherens junction marker AJM-1::GFP/MH27 antibody staining) and cells fuse together (Stages 0-2). Fusion progresses from the anterior to posterior of the tail tip. EM analysis of fusing cells suggest that fusions initiate at or near adherens junctions (Nguyen et al., 1999). |
|
MaleHypFIG 10: Tail tip hyp remodeling in the male. Note, hyp 8 (1 nucleus), hyp 9 (1 nucleus), hyp 10 (2 nuclei) and hyp 13 (2 nuclei) are included in the ventral hypodermal tissue (beige) in stages 0-4. Hyp 11 nucleus is omitted from these stages as it usually lies on the left lateral side, rather than ventral. In stage 5, because of the change of the tail tip hyp into a "central/dorsal hyp", hyp 11 nucleus is now also included.
The newly formed tail tip syncytium then migrates anteriorly into the body at the midline until it forms a strip of central hyp that extends from the tail to the anterior edge of the proctodeum (Stages 3-5 and adult in MaleHypFIG 11A&B).
MaleHypFIG 11: EM cross-section through the proctodeum.
3 List of Male Hypodermal Cells
(For cell name/number correspondence see HypFIG 1, HypFIG 2 in Hermaphrodite Hypodermis chapter. Dorsal nuclei locations are post-intercalation: LL: lateral left; LR: lateral right. V1/2, D1/2 etc indicate stochastic fates. Note, hyp 6 cells eventually fuse with hyp 7 during mid-L3 stage)
4 References
Baird, S.E., Fitch, D.H., Kassem, I.A. and Emmons, S.W. 1991. Pattern formation in the nematode epidermis: determination of the arrangement of peripheral sense organs in the C. elegans male tail. Development 113: 515-26. Article
Chow, K.L., Hall, D.H. and Emmons, S.W. 1995. The mab-21 gene of Caenorhabditis elegans encodes a novel protein required for choice of alternate cell fates. Development 121: 3615-26. Article
Dalpé, G., Zhang, L.W. Zheng, H. and Culotti, J.G. 2004. Conversion of cell movement responses to Semaphorin-1 and Plexin-1 from attraction to repulsion by lowered levels of specific RAC GTPases in C. elegans. Development 131: 2073-88. Article
Ginzburg, V.E., Roy, P.J. and Culotti, J.G. 2002. Semaphorin 1a and semaphorin 1b are required for correct epidermal cell positioning and adhesion during morphogenesis in C. elegans. Development 129: 2065-78. Article
Hahn, A.C. and Emmons, S.W. 2003. The roles of an ephrin and a semaphorin in patterning cell-cell contacts in C. elegans sensory organ development. Dev. Biol. 256: 379-88. Article
Nguyen, C.Q., Hall, D.H., Yang, Y. and Fitch, D.H.A. 1999. Morphogenesis of the Caenorhabditis elegans male tail tip. Dev. Biol. 207: 86-106. Article
Portman, D.S. and Emmons, S.W. 2000. The basic helix-loop helix transcription factors LIN-32 and HLH-2 function together in multiple steps of a C. elegans neuronal sublineage. Development 127: 5415-26. Article
Ross, J.M., Kalis, A.K., Murphy, M.W. and Zarkower, D. 2005. The DM domain protein MAB-3 promotes sex-specific neurogenesis in C. elegans by regulating bHLH proteins. Dev. Cell 8: 881-92. Article
Roy, P.J., Zheng, H, Warren, C.E. and Culotti, J.G. 2000. mab-20 encodes Semaphorin-2a and is required to prevent ectopic cell contacts during epidermal morphogenesis in Caenorhabditis elegans. Development 127: 755-67. Article
Sulston, J.E. and Horvitz, H. R. 1977. Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Dev. Biol. 56: 110-156. Article
Sulston, J.E., Albertson, D.G. and Thomson, J.N. 1980. The Caenorhabditis elegans male: Postembryonic development of nongonadal structures. Dev Biol. 78: 542-576. Article
Sulston, J.E., Schierenberg, E., White J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64-119. Article
Yi, W., Ross, J.W. and Zarkower, D. 2000. mab-3 is a direct tra-1 target gene regulating diverse aspects of C. elegans male sexual development and behavior. Development 127: 4469-80. Article
Zhao, C. and Emmons, S.W. 1995. A transcription factor controlling development of peripheral sense organs in C. elegans. Nature 373: 74-78. Abstract
|

This chapter should be cited as: Lints, R. and Hall, D.H. 2009. Male epithelial system, hypodermis. In WormAtlas. doi:10.3908/wormatlas.2.7
Edited for the web by Laura A. Herndon. Last revision: June 17, 2013. |
|