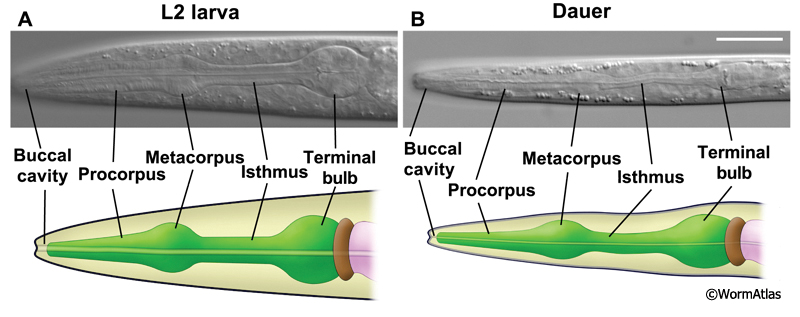
3 Pharyngeal Muscles
Eight sets of pharyngeal muscles (pm1-pm8), totaling 34 muscle cells, encircle the pharyngeal lumen. In non-dauers, these muscles contract in a coordinated fashion between 150-250 times/minute to concentrate and transport food particles to the grinder. Pharyngeal muscles contract radially, so that muscle contraction widens the central lumen. Relaxation of the pharyngeal muscles closes the lumen.
The pharyngeal muscles in the dauer were previously described to remain intact with abundant myofibrils (Popham and Webster, 1978). These authors also noted the presence of lipid and glycogen droplets and “large, elliptical mitochondria with electron-dense matrices”. Glycogen was proposed to provide a readily mobilized energy source for pharyngeal muscle contraction in response to a food source. The presence of mitochondria with electron-dense matrices was noted to be similar to mitochondria found in cells under limited substrate conditions, and possibly reflected the dauer’s non-feeding status. Although these muscles are shrunken in volume compared to L2 pharynx, they appear less drastically shrunken than the neighboring marginal cells.
3.1 Anterior pharyngeal muscles
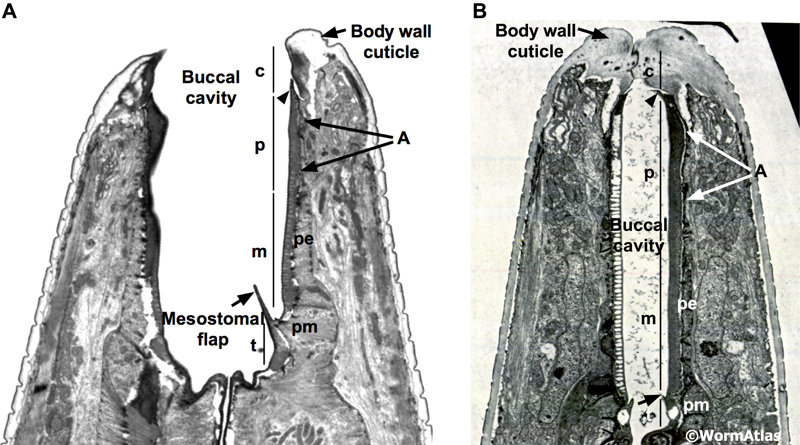
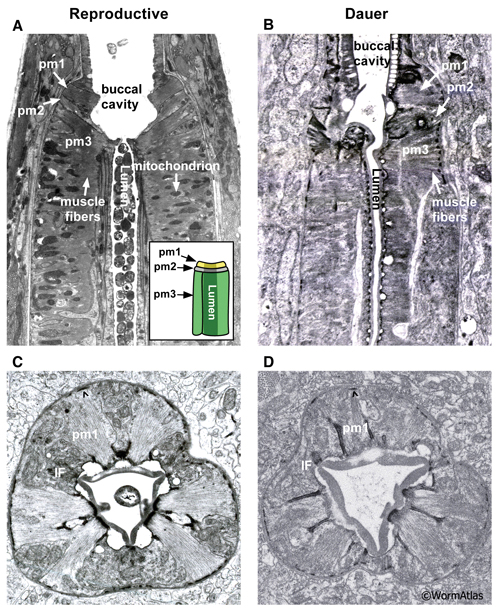
The anterior pharyngeal muscles, pm1 and pm2, are arranged into three groups. pm1 is the anteriormost pharyngeal muscle, forming a ring emerging behind the pharyngeal epithelial cells. The pm2 muscles also form a cylinder stacked behind pm1. Both the pm1 and pm2 cylinders are formed by cytoplasmic projections from cell bodies located more posteriorly along the pharynx. The transition between the pharyngeal epithelium and pm1 is preserved in the dauer and appears roughly similar to that in non-dauers (DPhaSUPFIG 1).
3.2 Procorpus and metacorpus muscles
The pm3 muscles are wedge-shaped, fitting together into a long cylinder that forms the procorpus. Cross-sectional EMs of the pharyngeal muscles in non-dauers reveal strips of single sarcomere muscle fibers projecting from the lumen to the basal lamina. In dauers, the strips of muscle fibers are narrow and shortened, suggesting a decrease in muscle fiber content (DPhaSUPFIG 2). The muscle cytoplasmic volume may also be reduced in dauers.
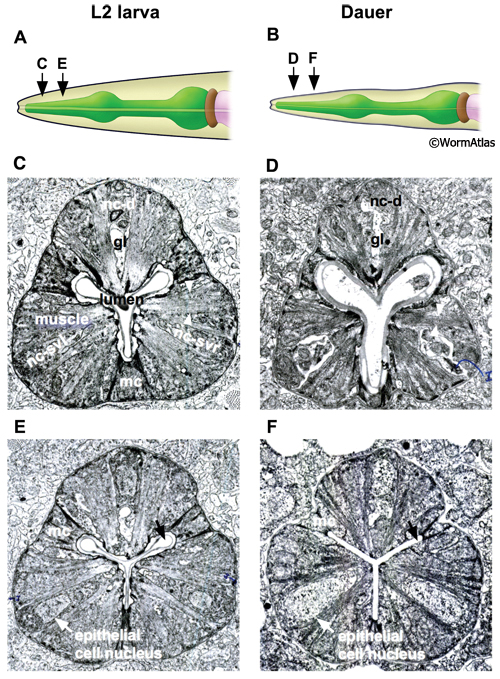
The pharyngeal lumen is the central canal through which food particles are filtered away from liquid and are passed into the terminal bulb, where a structure called the grinder crushes the food before passage to the intestine. In non-dauers, the lumen of the anterior and medial procorpus appears a three-pointed shape with tubular apices (DPhaSUPFIG 2) (Albertson and Thomson, 1975). Contraction of the pharyngeal muscles widens the lumen, allowing the passage of material, while pharynx muscle relaxation closes the lumenal opening except at each tubular apex. In dauers, the lumen of the medial procorpus is often closed, possibly indicating constitutive pharyngeal muscle relaxation in this region. Along with closure of the pharyngeal lumen in some dauers, the tubular apices are often closed (DPhaSUPFIG 2F). In the metacorpus and isthmus regions of the pharynx, the lumen is generally closed in both dauers and non-dauers (DPhaFIG 5&6B).
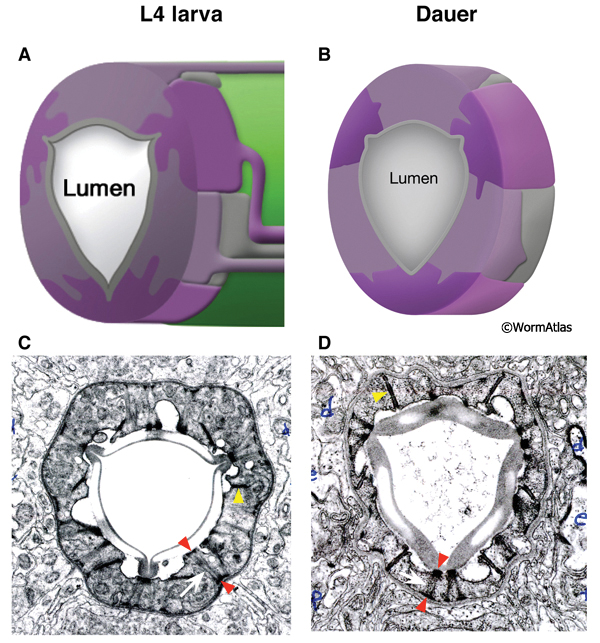
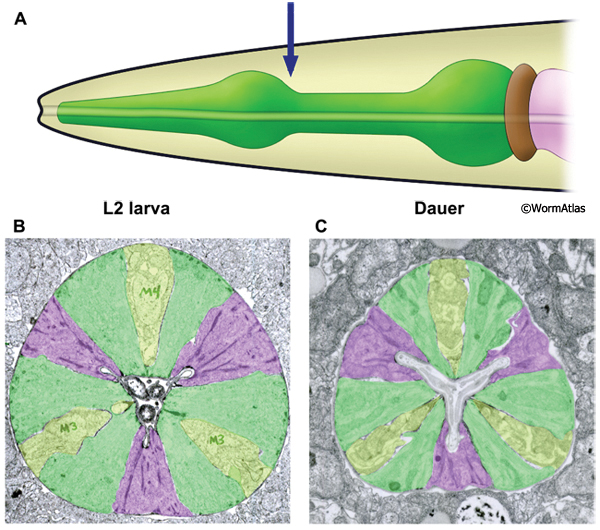
The first pharyngeal bulb, also named the metacorpus, is formed by the pm4 muscles (DPhaFIG 5, DPhaSUPFIG 3). The lateral borders between the muscle and marginal cells are reinforced by adherens junctions. Hemi-desmosomes anchor muscle fibers to the apical and basal membranes of each muscle cell. Shrinkage of the metacorpus is observed in dauers. Shrinkage appears to affect both the marginal cells and the muscles. The degree of shrinkage may affect the overall pharynx shape and appears to be somewhat variable between individual dauers.
DPhaFIG 5: Shape and musculature of dauer anterior metacorpus. A. Illustration of pharynx with arrow showing approximate position of transverse TEM sections in B&C. (Image source: WormAtlas PhaFIG 2A.) B. Transverse TEM section of L2 larva anterior metacorpus. Features of metacorpus are as follows: mc, marginal cell; nc-d, dorsal nerve cord; nc-svl, left subventral nerve cord; nc-svr, right subventral nerve cord. (Image source: N2 L2 28-14 [D. Riddle] 750.) C. Transverse TEM section of dauer anterior metacorpus. Arrowheads indicate muscle fibers for comparison in L2 and dauer images. Note heterogeneity in cross-sectional shape of dauer pharynx, less round than in the L2 larva. Images are not presented at the same scale, and were differentially magnified to show structural features. (Image source: N2 starved dauer 50-2-2 [D. Riddle] 462M.)
3.3 Isthmus muscles
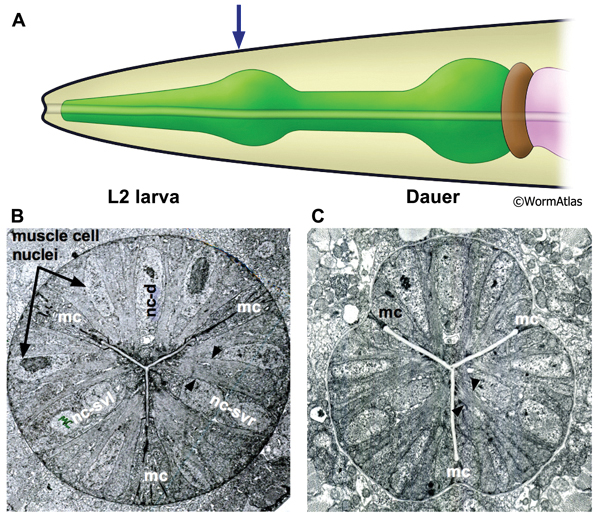
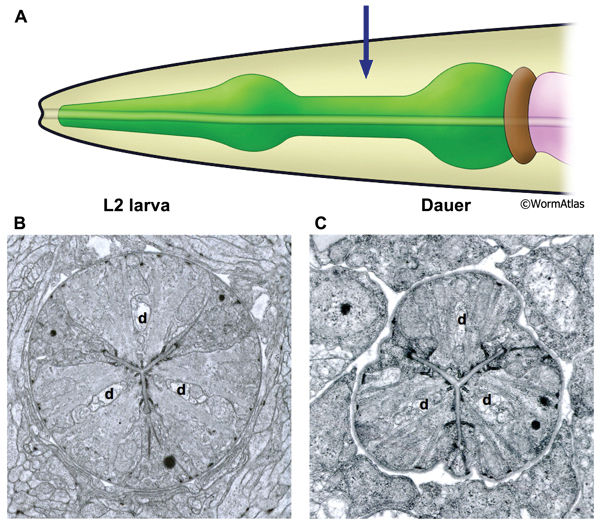
The isthmus is a narrowed region of the pharynx bridging the metacorpus and the terminal bulb. Three wedge-shaped pm5 muscles are arranged cylindrically around the central lumen to create the isthmus. Marginal cells join the pm5 cells at the apical margins. In non-dauers, the cylindrical isthmus appears circular in cross-section. In dauers, shrinkage of the marginal cells gives the isthmus a triangulated shape in cross-section (DPhaFIG 6). Despite the shrunken appearance of the isthmus, muscle fibers within the pm5 cells are still visible, but appear to be shortened. Individual bundles of actomyosin include fewer filaments.
DPhaFIG 6: Isthmus region of dauer pharynx. A. Illustration of pharynx with arrow indicating location of TEM sections. B. Transverse TEM section of pharynx in L2 larva in the region of the somatic nerve ring. (Image source: N2 L2 28-14 [D. Riddle] 1089.) C. Transverse TEM section of dauer larva in the region of the somatic nerve ring. (Image source: N2 starved dauer 50-2-2 [D. Riddle] 804.) Gland cell ducts are present in each pharynx region (d). Images are not presented at the same scale, and were differentially magnified to show structural features.
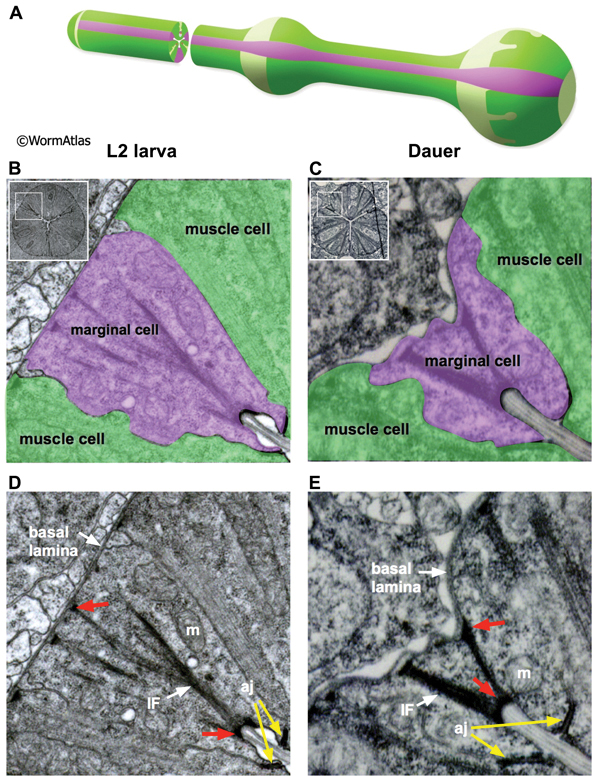
4 Marginal Cells
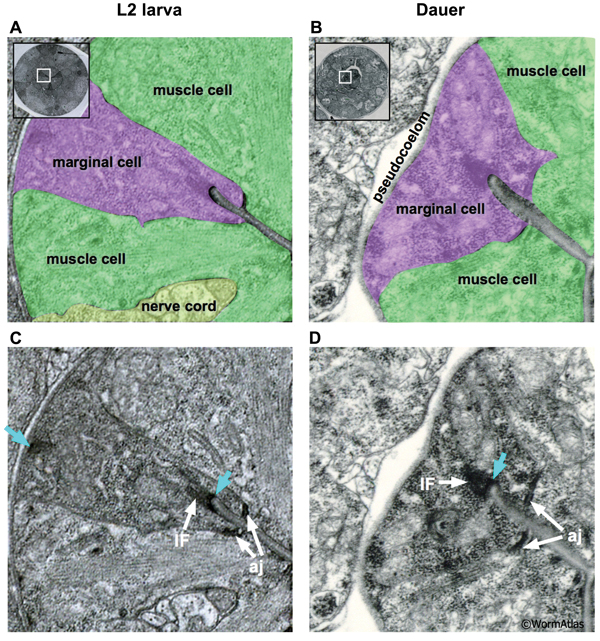
The C. elegans pharynx contains three types of marginal cells, named mc1-3. Each set of 3 marginal cells is situated at the apices of the muscle segments running along the length of the pharynx (See Pharynx section). In dauers, the marginal cells display variable morphology through the pharyngeal regions. In the procorpus and isthmus regions, the marginal cells are highly shrunken, giving the dauer pharynx a pinched appearance (DPhaFIG 7 and DPhaSUPFIG 4). In the metacorpus, each marginal cell must expand locally in size to accommodate the cell nucleus. The dauer marginal cells within the procorpus contain dense intermediate filaments (IF) bridging their apical and basal membranes. Studies of marginal cell structure in IF protein mutants may help to determine whether the IFs provide structural support to the dauer marginal cells (Karabinos et al., 2003). In contrast, IFs are relatively scarce in the isthmus (DPhaSUPFIG 4). Dauer marginal cells contain very thick adherens junctions in most pharynx regions. Mitochondria are abundant in the dauer marginal cells.
5 Gland Cells
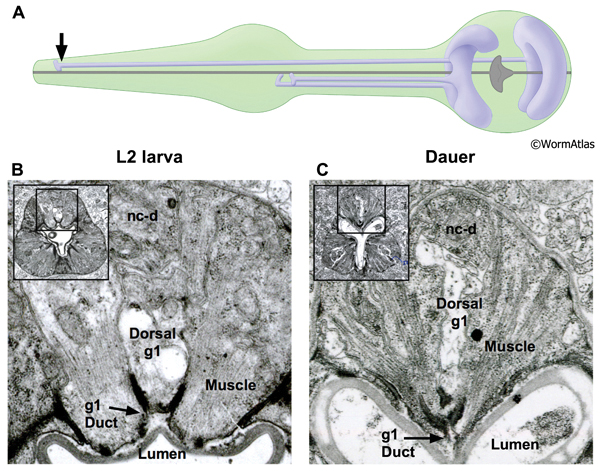
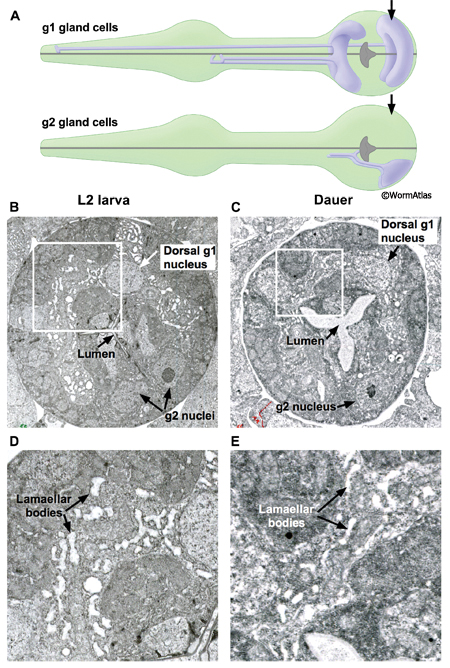
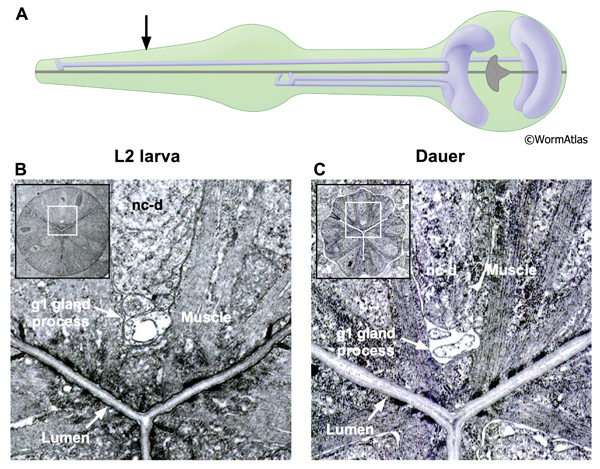
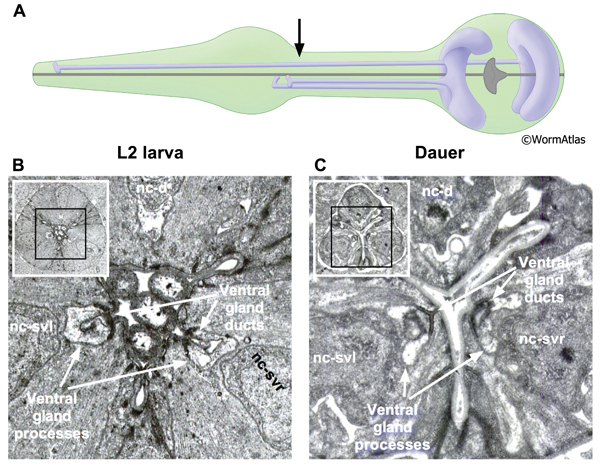
The pharynx contains two classes of gland cells, named g1 and g2. The gland cell bodies are located within the terminal bulb (PhaFIG 8A-G, PhaFIG 8H, PhaMOVIE 1). The g1 cell bodies contain numerous lamellar structures while the g2 gland cell bodies are clear and may contain vesicles. All five gland cells extend processes anteriorly into the isthmus along the dorsal and two subventral nerve cords. Thus, the three pharyngeal “nerve cords” encompass both neuronal and non-neuronal cells, without intervening basement membranes to separate the different cell types from one another. The processes of the subventral g1 glands and the g2 glands travel within the left and right subventral nerve cords to the metacorpus. In contrast, the dorsal g1 gland process follows the dorsal nerve cord and terminates in the procorpus. The gland cell processes open into the pharyngeal lumen at cuticle-lined ducts. The duct for the dorsal g1 is located in the anterior pharynx, posterior to the buccal cavity. The ducts for the left and right ventral g1 cells and for both g2 cells are situated to the posterior of the metacorpus.
In dauers, the g1 ducts, process and cell bodies are identifiable at the same positions as in non-dauers. The cuticle-lined gland cell ducts are still detectable in dauers. The most noticeable change is that the cross-sectional cytoplasmic volume of the g1 gland processes appears somewhat shrunken in dauers, possibly indicating a reduction in flow through these structures (DPhaFIG 8, DPhaSUPFIG 5 and DPhaSUPFIG 6). Shrinkage of the g1 gland processes was also observed in wildtype starvation-induced dauers as compared with daf-9(e1406) mutants, which form incomplete dauers without pharyngeal remodeling (Albert and Riddle, 1988). Similarly, the cytoplasmic lamellar bodies in the dorsal g1 gland cell body of a dauer larva appear collapsed in comparison to the same structures in non-dauers (DPhaFIG 9).
The function of these glands may be to aid either in digestion or molting, by providing enzymes packaged into vesicles to the pharyngeal lumen (Alberston and Thomson 1975). Secretion of g1 gland cell contents has been observed during feeding and molting in non-dauer larvae (Singh and Sulston, 1978; Sulston et al., 1983; Hall and Hedgecock, 1991). In non-dauer larvae, dark-staining granules appear in the g1 gland cells prior to the molt, which are thought to assist in the molting process (Singh and Sulston, 1978). Such granules were not observed in g1 glands of wildtype or daf-9mutant dauers (Albert and Riddle, 1988).
DPhaFIG 8: Dorsal g1 gland cell duct. A. Illustration showing positions of dorsal and two ventral g1 gland cells in pharynx. Arrow indicates approximate position of sections in lower panels. B. Transverse TEM section of anterior pharynx in L2 showing dorsal g1 gland (g1) in the vicinity of the cuticle-lined duct joining the gland process to the pharynx lumen. Insets show position of enlarged views on intact pharynx images. The dorsal nerve cord is indicated (nc-d). (Image source: N2 L2 28-14 [D. Riddle] 163.) C. Transverse TEM section from dauer larva showing duct of dorsal g1 gland cell. (Image source: N2 starved dauer 50-6-2 [D. Riddle] 293.)
DPhaFIG 9: G1 and g2 gland cells in dauers. A. Illustrations show the g1 and g2 gland cells in the pharynx. Approximate positions of TEM sections are indicated by arrows. B. Transverse TEM section of L2 pharynx terminal bulb showing the dorsal g1 and g2 cells. White box outlines the region enlarged in D. (Image source: N2 L2 28-14 [D. Riddle] 1502.) C. Transverse TEM section of dauer pharynx terminal bulb. White box outlines the region enlarged in E. (Image source: N2 daf-4(m63) dauer [D. Riddle] 227.) B&C. Arrows indicate nuclei of dorsal g1 and g2 cells. The extent of the dorsal g1 gland cytoplasm is easily seen by the swollen lamellae inside, while the g2 gland cytoplasm lacks such swollen lamellae. The extent of g1 lamellar swelling is much reduced in the dauer.
6 Pharyngeal Neurons
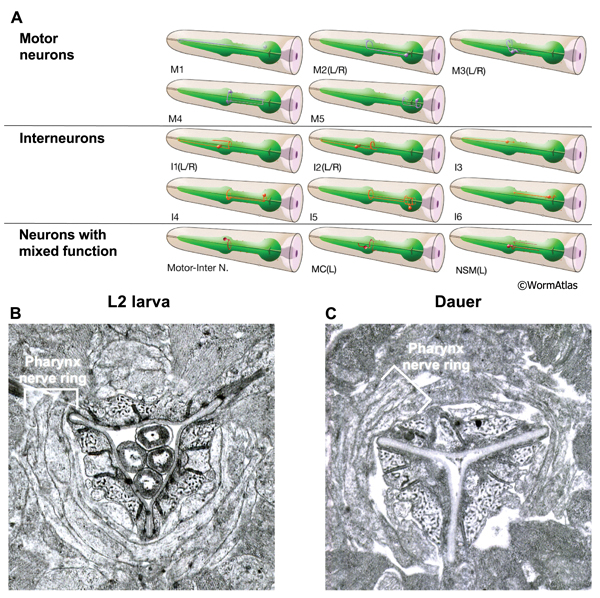
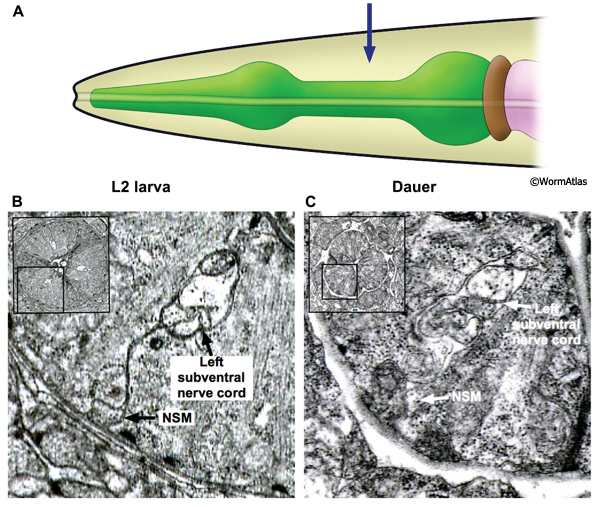
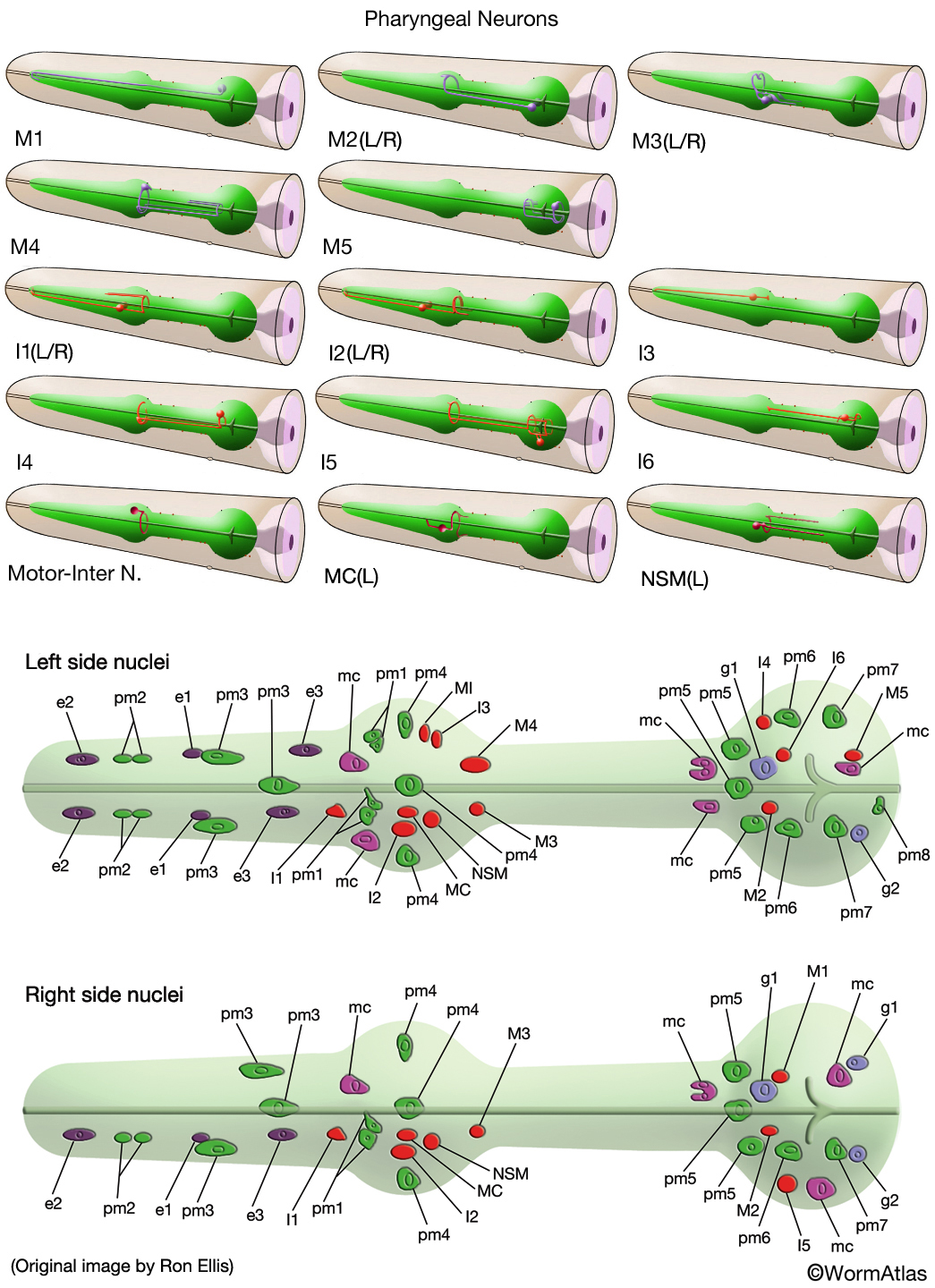
The nervous system of the pharynx consists of motor neurons, interneurons and polymodal neurons with mixed functions. The cell bodies of the neurons are located in two regions. Some cell bodies are located medially in the pharynx, within the posterior third of the procorpus and metacorpus. Other neurons have cell bodies located in the terminal bulb. The processes of the pharyngeal neurons primarily extend through the isthmus or the procorpus and are contained within one dorsal and two subventral nerve cords. In addition, the processes of some pharyngeal neurons decussate within the metacorpus to form the pharynx nerve ring.
In the dauer, the overall structure of the pharyngeal nervous system is maintained intact. Neuronal processes are approximately correctly positioned (DPhaFIG 10 and DPhaSUPFIG 7). The dauer pharyngeal neurons are not as shrunken as the pharyngeal epithelium and marginal cells (DPhaSUPFIG 8). The structure and function of pharyngeal neurons in the dauer have not been fully characterized and many aspects remain open to investigation.
DPhaFIG 10: Neuronal processes within the pharyngeal nerve ring. A. Illustrations of the pharyngeal neurons. Several pharyngeal neurons extend processes that decussate in the metacorpus to form a pharyngeal nerve ring. (Image source: WormAtlas.) B&C. The nerve ring is roughly intact in dauers, as compared to a similar region in an L2. There may be a loss of adherence between neuronal and extraneuronal tissues in the dauer, reflected by gaps between the dauer tissues. The pharyngeal pm4 muscles contain dark-staining tubules which are visible around the lumen in L2 and dauer larvae. B. Transverse TEM section of metacorpus at region of pharyngeal nerve ring in an L2 larva. (Image source: N2 L2 28-14 [D. Riddle] 803.) C. Transverse TEM section of metacorpus and pharyngeal nerve ring in a dauer larva. (Image source: N2 starved dauer 50-2-1 [D. Riddle] 316M.)
7 Terminal Bulb and Grinder
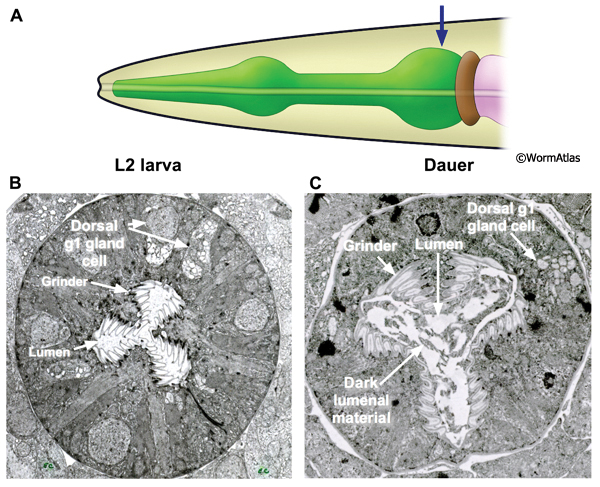
The terminal bulb (aka the posterior bulb, or 2nd bulb) of the pharynx contains about a dozen cell bodies of specialized muscles, neurons, gland cells and marginal cells (See PhaFIG 1). The terminal bulb directly abuts the six cells of the pharyngeal valve, which firmly links the pharynx to the intestine. During formation of the dauer larva, most tissues in the terminal bulb undergo substantial shrinkage in concert with the slimming of the rest of the pharynx. Only the pharyngeal neuron cell bodies seem to resist shrinkage. Three types of pharyngeal muscles lie within the terminal bulb, pm6, pm7 and pm8. The apical sides of the pm6 and pm7 muscles extend specialized tooth-like ridges that meet at the lumen to form the grinder (DPhaFIG 11). Radial muscle filaments inside pm6 and pm7 are fastened to these teeth, which are covered by a thickened cuticle secreted by pm6 and pm7. The pm8 muscle is a single cell that may squeeze the pharyngeal valve, possibly to control its lumenal diameter.
The grinder in the terminal bulb is a cuticular structure which macerates the animal’s food. The grinder may also act as a gate preventing backflow of pharyngeal or intestinal contents. The grinder in the dauer looks much like that in the L2 with two notable features (DPhaFIG 11). First, the dauer grinder occupies a larger proportion of the overall terminal bulb due to shrinkage of the terminal bulb’s cellular structures. Second, the lumen of dauer specimens often contains a dark-stained material not seen in the pharyngeal lumens of L2 larvae. The dark lumenal material may reflect incomplete shedding of the L2d pharyngeal cuticle. During molting, the pharyngeal cuticle is removed by digestion through the alimentary tract. Since dauers do not pump the pharynx, this mechanism for eliminating the molted pharyngeal cuticle could be impaired.
DPhaFIG 11: The terminal bulb and grinder. A. Illustration of the head and pharynx with region of TEM section designated by an arrow. B. Transverse TEM section of an L2 larva showing the pharyngeal terminal bulb. The locations of the grinder, pharyngeal lumen and dorsal g1 gland cell body are indicated. (Image source: N2 L2 28-14 [D. Riddle] 1422.) C. Transverse TEM section of a dauer larva showing the pharynx terminal bulb. The positions of the grinder, pharyngeal lumen and dorsal g1 gland cell are indicated. (Image source: N2 starved dauer 50-2-1 [D. Riddle] 973.) The overall structures of the terminal bulb and grinder are preserved in dauers. The grinder appears to occupy a larger relative portion of the terminal bulb area in the dauer, probably reflecting that the extracellular cuticle of the grinder teeth is virtually unchanged, while most of the terminal bulb's cells have shrunken dramatically. The L2 and dauer images are not to the same scale. In dauers, the pharyngeal lumen contains dark material which may be unabsorbed L2 cuticle.
8 Supplemental figures
DPhaSUPFIG 1: Anterior pharynx region. A&C. Anterior pharyngeal region of reproductively-developing animals. B&D. Anterior pharyngeal region of larvae arrested in dauer. A. Anterior pharynx of an adult (horizontal TEM section). (Image source: N533 [D. Hall] J2 F300.) Inset, illustration of anterior pharynx muscles, showing the stacked-cylinder arrangement of pm1, pm2 and pm3. Only two pm3 cells are shown; the third pm3 cell is behind the lumen out of the plane of view. The lumen is depicted as a central cylinder. When the pharynx muscles are relaxed, the lumen is closed. B. Horizontal TEM section of anterior pharynx in a dauer. (Image source: Starved N2 dauer 50-7-1 [D. Riddle] 34.) C. Anterior pharynx of an L2 (transverse TEM section). (Image source: N2 L2 28-14 [D. Riddle] 144.) D. Anterior pharynx of a dauer (transverse TEM section). (Image source: Starved N2 dauer 50-2-2 [D. Riddle] 225.) Karets indicate hemi-desmosomes anchoring pm1 muscle fibers to the basal lamina. Cytoplasmic intermediate filaments (IF) can be detected in the pharyngeal epithelial cell cytoplasm adjacent to the pm1 cells. The lumen of the nondauer animals contain ingested bacterial cells as evidence of feeding.
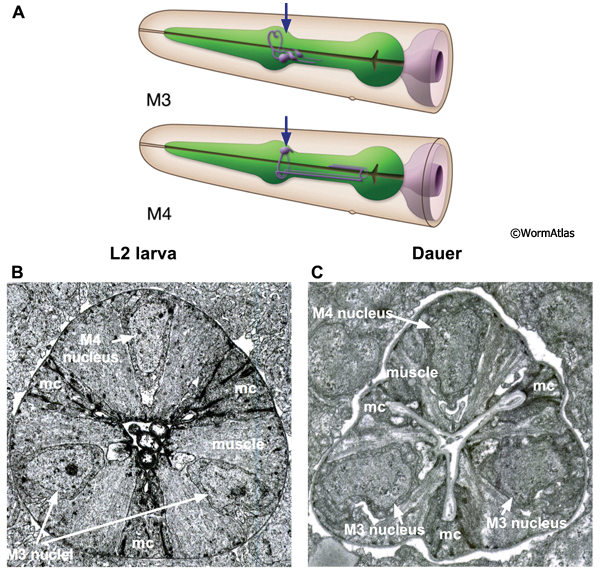
DPhaSUPFIG 3: Posterior metacorpus region of dauer pharynx. A. Illustration of pharynx with arrow to indicate location of TEM sections. B. Transverse TEM section of posterior metacorpus in an L2 larva. (Image source: N2 L2 28-14 [D. Riddle] 801.) C. Transverse TEM section of posterior metacorpus of a dauer pharynx. (Image source: N2 starved dauer 50-2-2 [D. Riddle] 556M.) B&C. Shading indicates cell types: green, muscle; purple, marginal cells; yellow, nerve cords containing neuronal and gland cells. Markings in B indicate M3 and M4 neuron cell bodies of three pharyngeal neurons in the metacorpus. Images are not presented at the same scale, and were differentially magnified to show structural features.
DPhaSUPFIG 4: Marginal cells in the isthmus region. A&C. Transverse TEM section from the isthmus region of the pharynx of an L2. A. Shading indicates cell types; muscles are green and marginal cells are purple. Inset shows location of enlarged region within cross-section of intact worm. C. Same section as in A, except without shading to show intracellular features. (Image source: N2 L2 28-14 [D. Riddle] 1147.) B&D. Transverse TEM section from the isthmus region of a dauer pharynx. B. Shaded as for A. D. Same section as in B, except without shading to show intracellular features. Blue arrows indicate hemi-desmosomes; IF, intermediate filaments; aj, adherens junctions joining marginal cells and neighboring muscle cells. L2 and dauer images are to different scales in order to increase visual resolution. (Image source: N2 starved dauer 50-2-1 [D. Riddle] 772a and 772 [inset in B].)
DPhaSUPFIG 5: g1 gland processes. A. lllustration showing positions of dorsal and ventral g1 gland cells. Arrow indicates approximate position of TEM sections in B&C. B. Transverse TEM section of L2 larva showing dorsal g1 gland process. (Image source: N2 L2 28-14 [D. Riddle] 750.) C. Transverse TEM section showing g1 gland process in a dauer larva. Contents of g1 process appears to be more empty and the process diameter is shrunken in the dauer compared to the L2 larva. Insets, boxed areas indicate regions in enlarged view; nc-d, dorsal pharyngeal nerve cord. (Image source: N2 starved dauer 50-2-1 [D. Riddle] 225M.)
DPhaSUPFIG 6: Ventral g1 gland ducts and processes. A. Illustrations showing positions of dorsal and ventral g1 gland cell bodies and processes. The gland processes travel anteriorly within the three pharyngeal nerve cords. The pharyngeal nerve cords contain neuronal and non-neuronal cell types in close apposition without intervening basement membranes. Arrow indicates approximate position of TEM sections in B&C. B. Transverse TEM section of L2 larva showing ventral g1 gland processes and ducts within the left and right subventral nerve cords. (Image source: N2 L2 28-14 [D. Riddle] 836.) C. Transverse TEM section of dauer showing ventral g1 gland processes within the left and right subventral nerve cords. Gland process diameters are shrunken in the dauer compared to L2. (Image source: N2 starved dauer 50-2-1 [D. Riddle] 350M.) Abbreviations: nc-d, dorsal nerve cord; nc-svl, left subventral nerve cord; nc-svr, right subventral nerve cord.
DPhaSUPFIG 7: Neuron processes in the isthmus. A. Illustration showing the head and pharynx with region of TEM sections designated by an arrow. B. Transverse TEM section in the isthmus of an L2 showing the left subventral nerve cord containing neuron processes and a gland cell process. The NSM process at the basal membrane of the pharynx is indicated. (Image source: N2 L2 28-14 [D. Riddle] 1035.) C. Transverse TEM section of a dauer pharynx in the isthmus region, also showing the left subventral nerve cord and an NSM process. (Image source: N2 starved dauer 50-2-2 [D. Riddle] 787.) In B&C, NSM processes in basal positions form active zones pointing towards the pseudocoelom (see also PhaFIG 9F-H to see example of these zones in the adult). This anatomical feature led to the hypothesis that NSM neurons have neurohumoral roles and may influence other cells at a distance (Avery and Thomas, 1997). One such example is the role of NSM serotonerigic neurotransmission in the enhanced slowing response of food-deprived animals upon encountering a food source (Sawin et a.l, 2000).
9 References
Albert, P.S. and Riddle, D.L. 1983. Developmental alterations in sensory neuroanatomy of the Caenorhabditis elegans dauer larva. J. Comp. Neurol. 219: 461-481. Abstract
Albert, P.S. and Riddle, D.L. 1988. Mutants of Caenorhabditis elegans that form dauer-like larvae. Dev. Biol. 126: 270-293. Abstract
Albertson, D.G. and Thomson, J.N. 1976. The pharynx of Caenorhabditis elegans. Phil. Trans. Royal Soc. London 275B: 299-325. Article
Altun, Z.F. and Hall, D.H. 2009. Alimentary system, pharynx. In WormAtlas. doi:10.3908/wormatlas.1.3
Avery, L. and Thomas, J.H. 1997. Feeding and defecation. In The nematode Caenorhabditis elegans (eds. D.L. Riddle, W.B. Wood). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. Article
Cassada, R.C. and Russell, R.L. 1975. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 46: 326-342. Abstract
Chow, D.K., Glenn, C.F., Johnston, J.L., Goldberg, I.G. and Wolkow, C.A. 2006. Sarcopenia in the Caenorhabditis elegans pharynx correlates with muscle contraction rate over lifespan. Exp. Gerontol. 41: 252-260. Abstract
Golden, J.W. and Riddle, D.L. 1984. The Caenorhabditis elegans dauer larva: Developmental effects of pheromone, food and temperature. Dev. Biol. 102: 368-378. Abstract
Hall, D.H and Hedgecock, E.M. 1991. Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans. Cell. 65: 837-847. Abstract
Karabinos, A., Schulz, E., Schünemann, J., Parry, D.A.D. and Weber, K. 2003. In vivo and in vitro evidence that four essential intermediate filament (IF) proteins A1, A2, A3 and B1 of the nematode Caenorhabditis elegans form an obligate heteropolymeric IF system. J. Mol. Biol. 333: 307-319. Abstract
Keane, J. and Avery, L. 2003. Mechanosensory inputs influence Caenorhabditis elegans pharyngeal activity via ivermectin sensitivity genes. Genetics. 164: 153-162. Article
Mango, S.E. The C. elegans pharynx: a model for organogenesis (January 22, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.129.1.
Meléndez, A., Tallóczy, Z., Seaman, M., Eskelinen, E. L., Hall, D. H., and Levine, B. 2003. Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science 301: 1387-1391. Abstract
Popham, J.D. and Webster, J.M. 1978. Aspects of the fine structure of the dauer larva of the nematode, Caenorhabditis elegans. Can. J. Zool. 57: 794-800. Abstract
Sawin, E.R., Ranganathan, R., and Horvitz, H.R. 2000. C. elegans locomotory rate is modulated by environment through a dopaminergic pathway by experience through a serotonergic pathway. Neuron. 26: 619-631. Article
Singh, R.N. and Sulston, J.E. 1978. Some observations on molting in Caenorhabditis elegans. Nematologica. 24: 63-71. Abstract
Sulston, J.E., Schlerenberg, E., White, J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64-119. Article
Wright, K.A. and Thomson, J.N. 1981. The buccal capsule of Caenorhabditis elegans (Nematoda: Rhabditoidea): an ultrastructural study. Can. J. Zool. 59: 1952-1961. Article
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image

{kind=link}