3 Locomotion
Dauer larvae have a tendency to remain motionless (Cassada and Russell, 1975; Gaglia and Kenyon, 2009) in either a rod-like or curved position (Gems et al., 1998, Lee et al., 2011). For a comparison of locomotion in dauers and L2/L3 larvae, see DBehaviorVID 1 and DBehaviorVID 2, respectively. Pharynx pumping also ceases in dauers. In these ways, dauer behavior is reminiscent of larval behaviors during the intermolt lethargus periods (Cassada and Russell, 1975). Mechanical stimuli, such as jarring the medium or touching the animal directly, induce an immediate and fast-paced locomotory response as the dauer flees the area (Cassada and Russell, 1975). This rapid locomotory response shows that the dauer’s body muscles are fully functional and that the tendency of dauers to lie motionless may reflect a shift in neuromuscular signaling, rather than a decline in muscle functional capacity. Curiously, conditions that alter the level of dopamine signaling (either up or down) can stimulate movement in dauer larvae, without altering baseline locomotion in adult animals (Gaglia and Kenyon, 2009). This evidence points to dopaminergic signaling as potential factor regulating locomotory behavior in the dauer.
DBehaviorVID 1: Dauer larvae locomotory behaviors. Dauer larvae on an agar surface lie motionless for much of the time. One larva moving through the field demonstrates the characteristic exaggerated sinusoidal behavior. (Video Source: Mark Alkema, University of Massachusetts Medical School.) To open high resolution video in a separate page click here. (Depending on connection speed, video may take several minutes to load.)
DBehaviorVID 2: Non-dauer larvae locomotory behaviors. In contrast to the motionless tendency of dauer larvae, non-dauer larvae are observed to move nearly constantly in the presence of food. Feeding larvae may or may not cover large distances when locomotory. Source: Mark Alkema, University of Massachusetts Medical School. To open high resolution video in a separate page, click here. (Depending on connection speed, video may take several minutes to load.)
4 Nictation
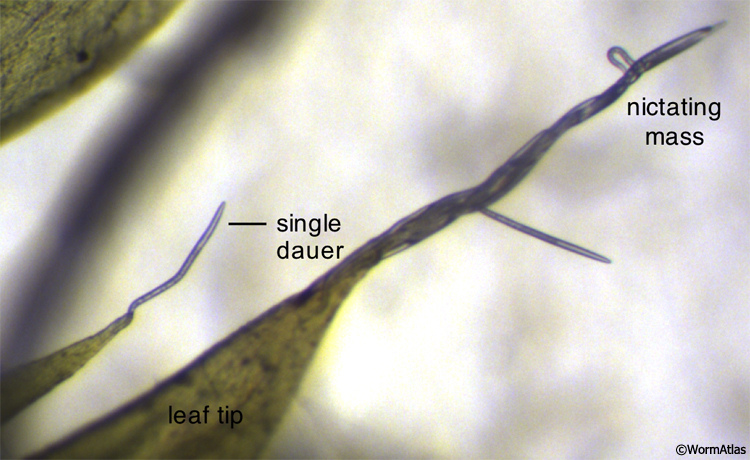
Dauers are sometimes observed performing a unique behavior called nictation. Nictating dauers have an erect posture, balancing on the end of their tails with their heads swaying in the air (Cassada and Russell, 1975) (DBehaviorFIG 1, DBehaviorVID 3, VID 4 and VID 5). Nictation behavior is specific to dauers and is not performed by directly-developing larvae or adults. On laboratory growth medium, C. elegans dauers are frequently found nictating upon filamentous structures, such as fungal hyphae, which commonly contaminate the agar growth medium. Dauers may also nictate as a mass, forming a structure resembling a waving stalk upon the substrate (DBehaviorVID 4) (Félix and Duveau, 2012). Controlled conditions for dauer nictation utilize artificial nictation substrates, such as medical cotton gauze and polydimethylsiloxane (PDMS)-based microfluidic structures (Lee et al., 2011).
DBehaviorFIG 1: Dauers performing nictation behavior. C. elegans dauers performing nictation behavior in the wild from the tips of pointed moss leaf tips. In the lower-left of the field, a single dauer is nictating alone. A large mass of dauers nictating together is shown in the center and right regions of the field. (Image source: Marie-Anne Félix, Institute of Biology of École Normale Supérieure.)
DBehaviorVID 3: Nictating C. elegans dauer larvae. Nictating dauers can be seen as objects waving upon filamentous nictation substrates, which may be debris or fungi accumulated on the agar surface. At the bottom-center of the viewer field, a group of dauers has become entwined together to form a larger nictating object. In addition, single dauers are nictating in other regions of the viewer field, including the left side. (Video source: Nathan Schroeder, Rutgers University.) Click "play" arrow to start video. To open high resolution video in a separate page click here. (Depending on connection speed, video may take several minutes to load.)
DBehaviorVID 4: A mass of thousands of dauers exhibiting nictation behavior. Dauers can be seen waving individually or in groups at the surface of any pointed structure. In this movie, thousands of dauers are observed oscillating en masse as found in the wild on a rotting apple (Félix and Duveau, 2012). This nictation behavior is thought to help in finding an invertebrate host for migration. (Video source: Marie-Anne Félix,Institute of Biology of École Normale Supérieure.) Click "play" arrow to start video. To open high resolution video in a separate page click here. (Depending on connection speed, video may take several minutes to load.)
DBehaviorVID 5: Dauers exhibiting nictation behavior along the top of an agar surface. In this movie, individual dauers are observed slowly waving back and forth. They appear to be standing on their tails and are waving their whole bodies in the air. The motion of the dauers is much slower compared to what is seen when they swimming in liquid and is different in waveform than when worms are crawling on a plate or swimming in liquid. Movie is in real time. (Video source: Mark Alkema, Universityof Massachusettes Medical School.) To open high resolution video in a separate page, click on video picture above or click here. (Depending on connection speed, video may take several minutes to load.)
Nictation requires cholinergic signaling within the ciliated IL2 neurons (Lee et al., 2011). In C. elegans dauers, the ciliated endings of the IL2 neurons are covered by a thick body wall cuticle over the animal’s nose (see DCutFIG 5), and the distal channel to the exterior is filled or narrowed with excess dense material (Albert and Riddle, 1981; DHH, unpublished observations), possibly interfering with transmission of chemical cues that might be related to nictation. Even in wild type adults, the IL2 cilia are somewhat difficult to dye-fill (Tong and Burglin, 2010), suggesting that the IL2 cilium is less exposed to the exterior than other chemosensors. Rather, IL2 neurons may initiate nictation in response to mechanosensory stimuli indicating appropriate environmental substrates for nictation (Lee et al., 2011). Nictation initiation is a dauer-specific function of IL2s, since activation of IL2s was not able to trigger nictation in non-dauers. In addition to mechanical stimulation of nictation, photostimulation with electric lights can trigger nictation behavior in starved populations of C. elegans (Staniland, 1957).
Nematodes demonstrate two strategies for foraging and dispersal. Nictation behavior exemplifies the ambushing, or “sit-and-wait”, strategy of host tracking and dispersal. The alternate dispersal mechanism utilizes cruising, or “widely-foraging”, in which nematodes actively explore their environment for a suitable food or hosts. While ambushing allows for targeting of quickly moving prey, (Huey and Pianka, 1981; Campbell and Gaugler, 1993), cruising is optimized for searching out sedentary, non-motile prey, such as rotting fruits.
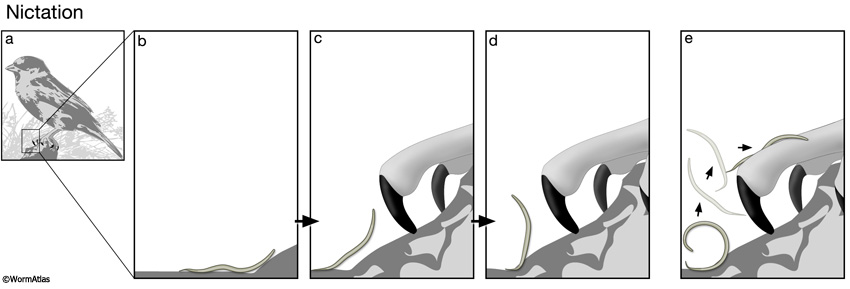
Nematodes in the Steinernema genus demonstrate an unusual variant of ambushing in which nictation is the first step of a more complex dispersal behavior. From a nictation position, an infective juvenile can bend backwards, progressively looping its body until the tail is flung away from the substrate, propelling the entire body towards the oncoming host (Campbell and Kaya, 1999; Hallem et al., 2011) (DBehaviorFIG 2). Jumping behavior in S. carpocapsae infective juveniles is stimulated by the presence of the nematode’s host, honeycomb moth larvae, but not by a non-specific cue, such as an air puff (Campbell and Kaya, 1999). Carbon dioxide gas can also stimulate S. carpocapsae jumping behavior in a BAG-dependent manner (Hallem et al., 2011).
DBehaviorFIG 2: Larval nictation and flipping behaviors for dispersal. A. Nematodes in the dauer or infective stages are adapted for dispersal to new environments in search of fresh food sources or for attaching to a new host, respectively. Both needs require mechanisms for attaching to a vector or host animal, such as a bird or insect. B-D. Larvae perform nictation behavior to increase their height above the surface, increasing the chances of being picked up by a passing object, such as the birds claw or an insects body. E. S. carpocapsae infective juveniles also perform jumping behavior to become airborne, thereby acquiring a more secure attachment to the passing animal.
5 Pharyngeal Pumping
The dauer animal has modified body structures to prevent normal feeding behavior, including a closed mouth and a somewhat withered pharynx. The pharyngeal muscles virtually cease pumping activity during dauer, although those muscles retain contractility. The pharyngeal muscles show altered responses to sensory cues, compared to other stages (Keane and Avery, 2003). In normal animals at any other larval stage or adulthood, light tail tap causes a brief halt (for several seconds) to the normal steady pumping of the pharynx when the animal is on food (Chalfie et al., 1985). A harsh tail tap in adults leads to a much longer period of inhibition, from which the animal only recovers slowly (Keane and Avery, 2003). Curiously, a harsh tail tap in dauers causes a brief stimulation of pharyngeal pumping (Keane and Avery, 2003). Both inhibitory and stimulatory responses are sensitive to mutations in unc-7, suggesting that gap junctions lie somewhere in the ill-defined behavioral pathway between the tail and the pharynx.
6 References
Ailion, M. and Thomas, J.H. 2000. Dauer formation induced by high temperatures in Caenorhabditis elegans. Genetics.156: 1047-67. Article
Albert, P.S. and Riddle, D.L. 1983. Developmental alterations in sensory neuroanatomy of the Caenorhabditis elegans dauer larva. J. Comp. Neurol. 219: 461-481. Abstract
Ashton, F.T., Li, J., and Schad, G.A. 1999. Chemo- and thermosensory neurons: structure and function in animal parasitic nematodes. Vet. Parasitol. 84: 297-316. Abstract
Bretscher, A.J., Busch, K.E., and de Bono, M. 2008. A carbon dioxide avoidance behavior is integrated with responses to ambient oxygen and food in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 105: 8044-8049. Article
Campbell, J.F. and Gaugler, R. 1993. Nictation behaviour and its ecological implications in the host search strategies of entomopathogenic nematodes (Heterorhabditidae and Steinernematidae). Behaviour 126:155-169. Abstract
Campbell, J.F. and Kaya, H.K. 1999. How and why a parasitic nematode jumps. Nature 397: 485-486. Abstract
Cassada, R.C. and Russell, R.L. 1975. The dauer larva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 46: 326-342. Abstract
Chalfie, M., Sulston, J.E., White, J.G., Southgate, E., Thomson, J.N. and Brenner, S. 1985. The neural circuit for touch sensitivity in Caenorhabditis elegans. J. Neurosci. 5: 956-964. Article
Chatzigeorgiou, M., Yoo, S., Watson, J.D., Lee, W.H., Spencer, W.C., Kindt, K.S., Hwang, S.W., Miller, D.M. 3rd, Treinin, M., Driscoll, M. and Schafer, W.R. 2010. Specific roles for DEG/ENaC and TRP channels in touch and thermosensation in C. elegans nociceptors. Nat. Neurosci. 13: 861-8. Article
Félix, M.-A. and Duveau, F. 2012. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biology 10: 59. Article
Gaglia, M.M. and Kenyon, C. 2009. Stimulation of movement in a quiescent, hibernation form of C. elegans by dopamine signaling. J. Neurosci. 29: 7302-7314. Article
Gems, D., Sutton, A.J., Sundermeyer, M.L., Albert, P.S., King, K.V., Edgley, M.L, Larsen, P.L., and Riddle, D.L. 1998. Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics 150: 129-155. Article
Guillermin, M.L., Castelletto, M.L., Hallem, E.A. 2011. Differentiation of carbon dioxide-sensing neurons in Caenorhabditis elegans requires the ETS-5 transcription factor. Genetics 189: 1327-1339. Abstract
Hall, D.H. and Treinin, M. 2011. How does morphology relate to function in sensory arbors? Trends Neurosci. 34: 443-51. Abstract
Hallem, E.A. and Sternberg, P.W. 2008. Acute carbon dioxide avoidance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. 105: 8038-8043. Article
Hallem, E.A., Dillman, A.R., Hong, A.V., Zhang, Y., Yano, J.M., DeMarco, S.F., and Sternberg, P.W. 2011. A sensory code for host seeking in parasitic nematodes. Curr. Biol. 21: 377-383. Article
Huey, R.B. and Pianka, E.R. 1981. Ecological consequences of foraging mode. Ecology 62: 991-999. Article
Keane, J. and Avery, L. 2003. Mechanosensory inputs influenceCaenorhabditis elegans pharyngeal activity via ivermectin sensitivity genes. Genetics 164: 153-162. Article
Lee, H., Choi, M., Lee, D., Kim, H., Hwang, H., Kim, H., Park, S., Paik, Y., and Lee, J. 2011. Nictation, a dispersal behavior of the nematode Caenorhabditis elegans, is regulated by IL2 neurons. Nature Neurosci. 15: 107-112. Abstract
Lopez, P.M., Boston, R., Ashton, F.T., and Schad, G.A. 2000. The neurons of class ALD mediate thermotaxis in the parasitic nematode, Strongyloides stercoralis. Int. J. Parasitol. 30: 1115-1121. Abstract
O’Halloran, D.M. and Burnell, A.M. 2003. An investigation of chemotaxis in the insect parasitic nematode Heterorhabditis bacteriophora. Parasitology 127: 375-385. Abstract
Procko, C., Lu, Y. and Shaham, S. 2011. Glia delimit shape changes of sensory neuron receptive endings in C. elegans. Development. 138:1371-1381. Article
Staniland, L.N. 1957. The swarming of Rhabditid eelworms in mushroom houses. Plant Pathol. 6: 61-62. Abstract
Tong, Y-G. and Burglin, T.R. 2010. Conditions for dye-filling of sensory neurons in Caenorhabditis elegans. J. Neurosci. Methods 188: 58-61. Abstract
Ward, S. 1973. Chemotaxis by the nematode Caenorhabditis elegans: Identification of attractants and analysis of the response by use of mutants. Proc. Nat. Acad. Sci. 70: 817-821. Article
Wittenburg, W. and R. Baumeister. 1999. Thermal avoidance in Caenorhabditis elegans: An approach to the study of nociception. PNAS 96: 10477-82. Article
Zhao, L.L., Wei, W., Kang, L., and Sun, J.H. 2007. Chemotaxis of the pinewood nematode, Bursaphelenchus xylophilus, to volatiles associated with host pine, Pinus massoniana, and its vector Monochamus alternatus. J. Chem. Ecol. 33: 1207-1216. Abstract
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image