The structure and connectivity of the
nervous system of the nematode Caenorhabditis elegans has been deduced from reconstructions of electron micrographs of serial sections. The hermaphrodite nervous system has a total complement of 302 neurons, which are arranged in an essentially invariant structure. Neurons with similar morphologies and connectivities have been grouped together into classes; there are 118 such classes. Neurons have simple morphologies with few, if any, branches. Processes from neurons run in defined positions within bundles of parallel processes,
synaptic connections being made en passant. Process bundles are arranged longitudinally and circumferentially and are often adjacent to ridges of hypodermis. Neurons are generally highly locally connected, making synaptic connections with many of their neighbours. Muscle cells have arms that run out to process bundles containing motoneuron axons. Here they receive their synaptic input in defined regions along the surface of the bundles, where motoneuron axons reside. Most of the morphologically identifiable synaptic connections in a typical animal are described. These consist of about 5000 chemical synapses, 2000 neuromuscular junctions and 600 gap junctions.
Introduction
The functional properties of a nervous system are largely determined by the characteristics of its component neurons and the pattern of synaptic connections between them. Although great progress has been made this century in understanding the manner in which information is coded within a neuron and the process of information transmission between neurons via synapses, little is currently known about the detailed connectivity of networks of neurons. The reason for this is simply that a nervous system is an enormously complex organ. In the vertebrate cerebellum alone, it has been estimated that there are more than 1010 neurons (Braitenberg & Atwood 1958) each making many thousands of synaptic contacts.
We have undertaken a complete reconstruction of a nervous system from electron micrographs of serial sections. We have been able to do this by using a very simple, small nervous system, that of the soil nematode Caenorhabditis elegans. The simplicity and consistency of structure of the nematode's nervous system attracted the attention of several neuroanatomists at the turn of the century. Richard Goldschmidt was perhaps the most notable of these; he attempted to reconstruct the nervous system of the large parasitic nematode Ascaris lumbricoides from serially sectioned material. Goldschmidt and his contemporaries produced detailed and accurate descriptions of the sensilla, the ganglia and the process tracts (Chitwood & Chitwood 1974), but the limited resolution of the light microscope prevented them from unambiguously resolving individual processes within bundles. Goldschmidt was convinced that neuron processes anastomosed extensively and that nervous tissue was therefore a syncytial network. He presented a set of intriguing diagrams representing the layout of processes in the Ascaris nervous system in support of his view of the structure of nervous tissue, a view that he vigorously defended (Goldschmidt 1908, 1909). The alternative viewpoint considered that neurons are mono nucleate branched structures and that their processes do not anastomose. It is now clear that this alternative viewpoint, as espoused by his contemporary critics, such as Cajal (1972), was correct. More recent anatomical studies with the electron microscope have finally laid to rest the reticularists' view of the nervous system. We have therefore not tried to interpret Goldschmidt's connectivity diagrams, although we have retained some of the names, given to the sensilla and ganglia, that were used by him and his contemporaries.
In recent years, C. elegans has become an object of intense developmental and genetical study. The highly reproducible sequence of cell divisions that takes place during the development of this organism has allowed the complete cell lineage to be determined from the fertilized zygote to the mature adult (Sulston 1983; Sulston et al. 1983). Each differentiated cell type that is produced at the terminal twigs on the lineage tree is now known. Laser ablation studies have given some insight into the degree of cell autonomy that is involved in determining the pattern of cell divisions and differentiations that occur. Generally it seems that, in C. elegans, cells behave fairly autonomously during development, although there are several well-defined instances where regulative cell-cell interactions have been demonstrated (Sulston & White 1980; Kimble 1981).
C. elegans was originally selected as an organism worthy of extensive developmental studies, partly because it is readily amenable to genetic analysis. Many mutants have been isolated and mapped (Brenner 1974). The mutants that have been isolated exhibit a wide variety of phenotypes: some are morphological, some affect various aspects of development and many exhibit aberrant behaviour. Some of the behavioural mutants have been shown to have defects in muscles (Waterston et al.1980), but many probably have alterations in the nervous system (Lewis & Hodgkin 1977; Chalfie & Sulston 1981; Hedgecock et al. 1984). It is hoped that a detailed knowledge of the structure of the wild-type nervous system of C. elegans will facilitate the interpretation of the changes that occur in such mutant nervous systems. This may in turn shed some light on the genetic control of the developmental processes that ultimately give rise to the specifically interconnected group of neurons that make up a nervous system.
The reconstructions that are presented in this paper describe the connectivity of all the neurons in the nervous system of the C. elegans hermaphrodite except those in the pharynx, which have been described by Albertson & Thomson (1976). The detailed morphologies of the sensilla in the head have been described by Ward et al. (1975), Ware et al. (1975) and Wright (1980); the structure of the ventral cord has been described by White et al. (1976) and an independent reconstruction of the tail ganglia has been described by Hall (1977). Together these papers give a fairly complete description of the connectivity, topography and ultrastructure of the nervous system in the hermaphrodite. The C. elegans male has a more extensive nervous system than that of the hermaphrodite; most of the 'extra' nervous tissue is situated in the tail. A partial reconstruction of the nervous system in the male tail has been described by Sulston et al. (1980).
The structure of the ventral cord of Ascaris has been deduced from reconstructions of light micrographs of serial sections (Stretton et al. 1978). In spite of the enormous difference in size between these two nematodes (10 cm as against 1 mm for C. elegans), the motoneurons in the ventral cord turn out to be remarkably similar, and it has been possible to identify equivalent motoneuron classes in the two animals. The large size of Ascaris enables electrophysiological techniques to be used in the study of its nervous system. Such studies have identified inhibitory and excitatory classes of motoneuron and have shown that acetylcholine is the neurotransmitter used by the excitatory motoneurons (Johnson & Stretton 1980). The small size of C. elegans precludes such electrophysiological studies but, by analogy, these results may be related to the equivalent neurons inC. elegans and so provide clues as to their functional properties.
Although reconstructions of nervous tissue from electron micrographs can in principle identify all focal synaptic contacts, it is unlikely that the pattern of connectivity obtained would exactly represent the functional synaptic connections between neurons. There is evidence that synaptic transmission mediated by some peptide transmitters acts over a considerable range (Jan et al.1983), suggesting that these types of synapses may not be localized at discrete focal contacts and therefore would not be seen in electron micrographs. There are other routes by which transmission of information could occur between neurons which are not apparent from reconstructions. Neurohumoral transmission is probably used for transmission over long distances and where many targets may be involved; a good candidate for a neurosecretory neuron has been found in the pharynx (Albertson & Thomson 1976). Short-range transmission may occur by means of electrical leakage currents or by capacitive coupling between processes that run alongside each other for long distances. However, in spite of these limitations, high-resolution reconstructions provide a wealth of information on the synaptic contacts between neurons. Thus, of all the currently available techniques, such reconstructions probably provide the most comprehensive picture of the synaptic circuits of a nervous system such as that of C. elegans.
Because of the large amount of information that is involved in presenting the connectivity data, we have tried to organize its presentation in such a way as to facilitate quick access. The structure of a 'canonical' nervous system is presented, which is in fact a mosaic of several nervous systems. A general description is first given of the structure of C. elegans and some of the salient features of the nervous system. This is followed by a detailed description of each of the neuron classes arranged in alphabetical order in Appendix 1. These descriptions are fairly self-contained and include morphological as well as synaptic data. There are many references in the first section to illustrations in Appendix 1. These appear as the class name followed by a letter, e.g. ASE-a. The lower case letter indicates the diagram referred to in the description of the neuron class ASE.
The reconstructed nervous systems described in this study were all derived from the nematode Caenorhabditis elegans (var. Bristol); these were cultured on lawns of E. coli grown on agar Petri plates (Brenner 1974).
Electron Microscopy
Worms were rinsed off Petri plates and fixed in 1% osmium tetroxide in 0.1 M sodium phosphate, pH 7.4 for one hour at 20 °C. Pre-fixing in glutaraldehyde was not done in this work because, although this method gives better preservation of fine structure, we found that osmium alone gave better contrast to cell membranes, and this facilitated the resolution of process outlines in regions of dense neuropile.
After fixation, the worms were spread on a thin layer of 1% agar and cut in half. The cut worms were covered with a drop of molten 1% agar, and blocks of agar containing a single half worm were cut out. These were dehydrated through a graded series of alcohols to propylene oxide, then to propylene oxide plus Araldite (CY 212 resin, CIBA Ltd.) and then into Araldite at room temperature overnight. The following day they were transferred to fresh Araldite and polymerized in gelatin capsules overnight at 60 °C.
An LKB ultratome III was used with a diamond knife to cut transverse serial sections of approximately 50 nm thickness. Ribbons of sections were generally picked up on Formvar coated 75-mesh copper grids. The sections in the region of the head, where most of the nervous system is situated, were picked up on slot grids, as it was found to be necessary to have every section in this region for successful reconstructions. Grids were stained with a 5% aqueous solution of uranyl acetate for 10 min at 60 °C and then with lead citrate for 5 min at room temperature according to the procedure of Reynolds (1963). Sections were photographed on cut film with an AEI 6B or an AEI 802 electron microscope. Most reconstructions were done directly from prints of micrographs of nervous tissue. In the region of the nerve ring, four-way montages were necessary; in other regions, single prints were sufficient. Every section was photographed in the region of the nerve ring and other areas of dense neuropile: photographs of every third section usually sufficed for following process bundles. Some use was made of a computer-aided reconstruction system described by White (1974) and Stevens & White (1979), but most of the reconstructions were done by hand from a total of about 8000 prints.
Small groups of processes were given arbitrary labels, which were written onto the prints with Rotring drafting pens. These labels were carried through all the pictures in which the associated processes were present, and this procedure was repeated until all process profiles were labelled. Processes could then be joined to other processes where branches had occurred, or ultimately be assigned to particular neurons if their cell bodies were within the scope of the reconstruction. When all the labelling was completed, each process was individually followed through every section in which it appeared, and a list was compiled of all the synaptic contacts that it made. In this way all synaptic contacts were recorded twice, once for each member of an interacting pair of processes. This provided a useful check on synapse scoring as any synaptic contact that was only scored once was reappraised.
The reconstructions were done piecemeal with data from five overlapping series; these were designated N2T, N2U, JSH, N2Y and JSE (figure A1, Appendix 1). The structure was found to be sufficiently invariant for equivalent processes and cell bodies to be identified in the region of overlap of two series. The N2T series was the first extended series to be cut in the head; the reconstructions of the head sensilla described by Ward et al. (1975) were based on this series. Although this series extended through the nerve ring and into the ventral cord, mesh grids were used and it was found that the inevitable occasional section loss, through obscuration by grid bars, allowed only a limited reconstruction to be done of these regions. The N2U series was from an old hermaphrodite that gave good quality pictures. It was sectioned on slot grids through the nerve ring and anterior ventral cord and a complete reconstruction of this region was obtained. This series also covered more than half the body length of the animal and enabled the anterior ventral and dorsal cords to be reconstructed. The JSH animal was a fourth stage (L4) larva, which was sectioned on slot grids. A complete reconstruction of the nervous system in the nerve ring and anterior ventral cord was obtained from this animal. This allowed the structure deduced from the N2U series to be validated in these regions, which are the most difficult to reconstruct because they contain dense neuropile with many processes that run close to the plane of sectioning. Few significant differences in structure that could be age-related were seen between the N2U and JSH series. The tail ganglia and some of the posterior ventral and dorsal cord were covered in the JSE reconstruction. The region between the anterior extremity of the JSE series and the posterior extremity of the N2U series has not been reconstructed in a hermaphrodite. A long series that overlapped at both ends, designated N2Y, was obtained from a male animal (Sulston et al.1980, in which it was referred to as series 4). The motoneurons of the ventral cord and the cells from the posterior lateral ganglion were reconstructed from this animal. The motoneurons (with the exception of the sex-specific VCn class) exhibited essentially the same synaptic behaviour as their anterior counterparts in the hermaphrodite. As there was also no reason to expect any sex-related differences in the cells of the posterior lateral ganglia, these data were incorporated to enable a complete reconstruction of the whole nervous system to be obtained. The structure that is described is a composite that has been derived from all these series except JSH.
Reliability of data
The biggest problem that was encountered in the course of the reconstruction work was the location of errors. Errors were generally made in one of three ways. (1) The most prevalent was human error, which would occur when following long featureless process bundles and which typically resulted in switches in process labels. (2) Many processes run close to the plane of sectioning in the vicinity of the nerve ring, with the result that the membranes of these processes would often be cut obliquely and give indistinct images. This made process identification very difficult in such situations, leading to the second most prevalent source of errors. (3) Similar errors of process identification also occurred in regions of poor image quality caused by dirt on sections or loss of sections on grid bars although, surprisingly, this was the least prevalent source of errors.
Errors generally manifested themselves by the appearance of an improbable structure, such as a process that was joined to more than one cell body or conversely not joined to any at all. Much of the nervous system was found to be bilaterally symmetrical; some of the sensory receptors in the head have higher levels of symmetry. Any deviations that were seen from expected symmetries were considered suspect. Errors were located either by exhaustive searching of every section that contained the process that was in question, or by looking at the reconstructions for discontinuities in synaptic behaviour, and then closely checking the regions of the process where the discontinuities occurred. In this way a complete, self-consistent structure was built up. The structures of the major regions of neuropile have been validated by separate reconstructions; the JSH series in the case of the nerve ring and the N2S series in the case of the ventral cord (White et al. 1976). Hall has undertaken an independent reconstruction of the tail ganglia; the structure that he describes is essentially the same as the structure that we describe here (Hall 1977).
We are reasonably confident that the structure that we present is substantially correct and gives a reasonable picture of the organization of the nervous system in a typical C. elegans hermaphrodite. It is likely that in the elaboration of a structure of this complexity that a few small errors might have crept in, but we feel that these may be quite limited because of the amount of cross-checking that was done. A few minor ambiguities still exist, however, which would require a considerable effort to clear up. These are described in Appendix 2.
Nomenclature
We have adopted a uniform system of nomenclature for naming the neurons and associated cells of C. elegans. Unfortunately it was not practicable to make such a system compatible with the various nomenclatures that have been used up till now. Appendix 3 lists the equivalences between these systems and the one used in this study. Neurons are given arbitrary names consisting of three upper case letters. The last letter can alternatively be a number of up to two digits. Additional symmetry descriptors are added to the name in the cases of groups of cells that are in the same class and related to each other by simple geometric symmetries. These descriptors are D or V (dorsal or ventral) and L or R (left or right). A group of cells with six-fold symmetry, such as IL1, has as its members: IL1DL, IL1DR, IL1L, IL1R, IL1VL and IL1VR. The members of the classes of motoneuron in the ventral cord do not have these symmetrical relations with each other. In these cases, the third digit of the class name is a numeral, which represents the anterior or posterior location of the neuron relative to its fellow class members; for example, VA3 is the third VA motoneuron. The use of the three-letter name without descriptors implies all members of the class if there is more than one. For the motoneurons, a lower case n is used in the third digit position to represent the generic name for all class members (for example, VAn). A slight modification of this system is used to describe the associated cells of sensilla, i.e. the sheath and socket cells. A sheath cell is designated by 'sh' and a socket cell 'so'. Thus in the case of the right sub-dorsal cephalic sensillum, the neuron is referred to as CEPDR, the sheath cell as CEPshDR and the socket cell as CEPsoDR.
Behaviour The animals pass through four larval stages before reaching adulthood: Ll, L2, L3 and L4. Each stage is terminated by a moult. If food is scarce, animals can go through an alternative developmental sequence in which a resistant 'dauer' larval form is produced at the L2 to L3 moult. Dauers can survive extreme conditions (desiccation and lack of food) for long periods until conditions improve and food becomes available, at which time they will moult and become normal adults (Cassada & Russell 1975; Riddle et al. 1981). Several structural changes occur on entering the dauer stage, including alterations to the endings of some sensory receptors (Albert & Riddle 1983).
C. elegans normally inhabits the interstices between damp soil particles or in rotting vegetation. It lives in a film of water and is held to solid surfaces by surface tension. Locomotion is achieved by dorso-ventral flexures of the body, which give rise to sinusoidal wave propagation along the length of the body. This can either be in the anterior-to-posterior direction, giving rise to forward motion, or in the posterior-to-anterior direction, giving backward motion. The head has an extra degree of freedom, in that it can make lateral as well as dorso-ventral movements. The dorso-ventral flexures (with the consequential sinusoidal posture of the body), combined with the surface tension forces, constrain the animals to lie on their sides. The L1, dauer and adult stages have longitudinal lateral ridges of cuticle, the alae, which may act to increase lateral friction and minimize sideslip. The thickness of the water film is quite critical; too thin or no water film results in the animals' becoming desiccated and dying, whereas if the film is greater than their diameter they are not held down to the surface and are unable to make any progress. C. elegans can move well on an agar surface even though this must be quite different from its normal habitat. If there is no food available locally it will move forward for quite long periods with occasional short intermissions of reversing. When it locates food it starts eating and stops moving, except for short foraging excursions forwards and backwards. Eggs tend to be laid only when the hermaphrodites have a plentiful food supply.
C. elegans responds in a regulated manner to a number of sensory stimuli: it will chemotax up a gradient of chemical attractant or down a gradient of repellant (Ward 1973; Dusenbery 1974); it will avoid regions of high osmolarity (Culotti & Russell 1978); it will actively maintain itself at an optimum temperature in a temperature gradient (Hedgecock & Russell 1975) and it will respond to light touch by moving away from the point of stimulation (Chalfie & Sulston 1981). In addition to these responses, the worm presumably uses its mechanosensory system to navigate through the interstices between soil particles in its natural habitat. Mating-specific behaviour is exhibited only by the male (Hodgkin 1983), which has additional neural circuitry in the tail for controlling copulation (Sulston et al. 1980).
Structure
The animal is ensheathed in a tough impermeable elastic cuticle, which is laid down by a system of underlying hypodermal cells. The body cavity (the pseudocoelome) is maintained at a high hydrostatic pressure relative to the outside; it is this pressure, acting on the elastic cuticle, which gives the animal its rigidity (the so-called hydrostatic skeleton (Crofton 1966).
There are four longitudinal ridges running down the inside of the body cavity: two medial and two lateral. These ridges consist of a ridge of hypodermis adjacent to a bundle of nerve processes, the whole structure being bounded by a basal lamina. Body movements are mediated by four strips of muscle cells running in four quadrants between these longitudinal ridges. Muscle cells have no obvious attachment points at either end and probably have attachments to the hypodermis distributed along their length. They act to deform the cuticle elastically against the stress produced by the turgor pressure.
Food is pumped into the animal and processed by a prominent pharynx. This is a virtually self-contained organ with its own musculature, epithelium and nervous system, and has been described in detail by Albertson & Thomson (1976). The pharynx probably functions as a largely autonomous unit, although there are two interneurons that originate in the central nervous system and enter it. These interneurons (RIP) are exclusively postsynaptic outside the pharnyx and so probably mediate the overall control of pharyngeal pumping from the central nervous system. The pharynx is used for ingesting food (usually bacteria), concentrating it by filtration and then grinding it, and probably also for secreting digestive enzymes from its gland cells (Albertson & Thomson 1976). The processed food is pumped into the intestine, which has a lumen lined with microvilli. The intestine is connected with the anus; defecation is controlled by three sets of specialized muscles (figure 12).
There is an excretory system, which consists of a single excretory canal cell arranged in an, 'H' configuration (Bird 1971).The two arms of the H run longitudinally down the lateral lines. These are joined by a cross bridge, which is connected to the excretory duct on the ventral side; this opens to the outside of the animal via the excretory pore situated on the ventral mid-line. Two ventrally situated 'gland' cells have anteriorly directed processes, which fuse and connect to the lumen of the excretory canal near the pore (Nelson et al. 1983). These processes continue running anteriorly on the ventral surface of the ventral nerve cord (figure 16) until the nerve ring is reached, where they terminate. The function of these glands is not yet known.
The adult hermaphrodite reproductive system consists of symmetrical pairs of uteri, oviducts, spermathecae and ovaries, which are joined at the uteri and connect to a vulva. This is situated on the ventral mid-line about halfway down the body (Hirsh et al.1976). During development, sperm are produced before oocytes and are stored for subsequent use. Egg-laying is mediated by a set of sixteen muscle cells, eight of which act to squeeze the contents of the uteri and eight to open the vulval orifice (figure 11).
The male gonad joins the rectum via the vas deferens to form a cloaca in the tail (Sulston et al. 1980). The cloaca is surrounded by a large, fan-like, copulatory bursa, which is richly endowed with sensory endings. These endings are derived from male-specific neurons, which are generated post-embryonically along with other neurons in the male. The male also has extra ventral body muscles and muscles that control the copulatory spicules (Sulston et al. 1980).
Organization of the nervous system and musculature
There are 302 neurons in the nervous system of C. elegans; this number is invariant between animals. Each neuron has a unique combination of properties, such as morphology, connectivity and position, so that every neuron may be given a unique label. Groups of neurons that differ from each other only in position have been assigned to classes. There are 118 classes that have been made using these criteria, the class sizes ranging from 1 to 13. Thus C. elegans has a rich variety of neuron types in spite of having only a small total complement of neurons. This is in marked contrast to structures such as the mammalian cerebellum, which contains more than 1010 neurons (Braitenberg & Atwood 1958) and yet has only five classes of component neuron (Eccles et al. 1967). Sensory transduction
The bulk of the nervous system of C. elegans is situated in the head, which is richly endowed with sensory receptors. These are arranged in groups of sense organs, known as sensilla. The arrangement and structure of sensilla have been described in detail (Ward et al. 1975; Ware et al. 1975; Wright 1980). Each sensillum contains one or a number of ciliated nerve endings and two non-neuronal cells: a sheath cell and a socket cell. A socket cell is effectively an interfacial hypodermal cell acting to join the sensillum to the hypodermis. A sheath cell is a glial-like cell that envelops the endings of neurons. Its inner surface, adjacent to the neural dendrite, is extensively invaginated and large number of secretory-like vesicles are often present in the cytoplasm. The sheath cells of the cephalic sensilla have, in addition, flat sheet-like processes that partly envelop the neuropile of the nerve ring and the anterior extremity of the ventral cord (figure 16). The function of sheath cells is not known, but they probably act to establish a defined extracellular milieu for the receptor endings.
Two large sensilla, the amphids, are located laterally and have internal channels, formed by the sheath and socket cells, which open through the cuticle to the outside. Eight neurons have their ciliated endings in this channel; a further four are associated with the sheath cell. There are two analogous structures, the phasmids, in the tail, but they are simpler in that they only have two neurons ending in the channel. The amphids and phasmids are generally considered to be the main chemoreceptive organs in the animal, because their structure permits a group of nerve endings to be exposed to the external environment of the animal.
The other sensilla in the head are arranged into two concentric rings around the mouth (figure 1). There is an inner ring of six, the inner labial sensilla, each of which has two associated neurones (IL1 & IL2). The dendrites of IL2 penetrate the cuticle to the outside of the animal and so they could be chemoreceptors. The other ending (ILl) lies embedded in the cuticle. There is an outer ring of four sensilla, the quadrant outer labials (OLQ), and these are paired with another set of four, the cephalic sensilla (CEP). Two additional lateral outer labial sensilla (OLL) are situated next to the amphid channel openings. The only other sensilla in the hermaphrodite are two pairs of lateral sensilla, the deirids, situated laterally in the anterior body (ADE) and the posterior body (PDE). These sensilla have similar morphologies to the cephalic sensilla in the head (Ward et al. 1975).
FIGURE 1. Sensory receptors in the head, as seen in an idealized section near the tip of the head. This region is richly endowed with sensory receptors, which are organized in a precise, complex arrangement. Most of the receptors are components of sensilla, and have associated sheath and socket cells. The amphid sensilla are situated in the lateral labia and have channels that are open to the outside with ADL, ADF, ASG, ASH, ASE, ASI, ASJ and ASK entering them. AWA, AWB, AWC and AFD are associated with the amphid sheath cells. There is a single inner labial sensillum in each labium, containing IL1 and IL2 receptor neurons. These sensilla also have channels to the outside, through which the processes of IL2 project. The two dorsal and the two ventral labia each have a single cephalic sensillum with a CEP receptor, and a single outer labial sensillum with an OLQ receptor. The lateral labia also each have an outer labial sensillum but with an OLL receptor. FLP and BAG are ciliated receptors that are free inside the head and are not part of a sensillum. URX and URY are not ciliated but have specialized flattened endings, which insinuate themselves around the inner and outer labial sensilla.
(See Figure 1 in color.)
In addition to the neurons of the sensilla there are other classes of neuron, which, on the basis of their connectivity and morphology, also probably serve a sensory transduction function. The best characterized neurons of this type are the touch receptors ALM, PLM, AVM and PVM. These have specialized, microtubule-filled processes, which run in close apposition to the hypodermis (Chalfie & Sulston 1981).
Disposition of cell bodies and ganglia
Several ganglia have been described and named in the nervous systems of other nematodes (Chitwood & Chitwood 1974). We have retained these names, where appropriate, for the ganglia in C. elegans. In several regions, cells are grouped together into well-defined ganglia by the arrangement of the basal lamina in the pseudocoelome. This sometimes results in adjacent cells' being partitioned into different ganglia. The lateral and ventral ganglia are not obviously separated in figure 2, for example, but in fact the cells of the ventral ganglion are a well-defined group (figure 3), being separated from those of the adjacent lateral ganglia by two basal laminae (figure 13). The arrangement of the basal lamina around the pseudocoelome will be discussed later; we will now describe the disposition of the various ganglia.
FIGURE 2. The locations of the cell bodies of all the neurons and their associated cells in the head is shown in left-hand (a) and right-hand (b) views. Cells marked with an asterisk are on or near the centre line and are shown in both views. These diagrams were derived from reconstructions of electron micrographs of one animal and, because of the difficulty of accurately measuring section thickness, there may be some longitudinal distortion. This is not excessive, however, as the overall longitudinal scale was normalized to views taken from the light microscope. The anterior bulb of the pharynx fits tightly in the body hypodermis and excludes cell bodies in the region of its maximum diameter. Cell bodies that are in this region are sometimes indeterminate as to which side of the bulb they reside, as in OLQsoDL/R. The neuropile of the nerve ring also excludes cell bodies and gives rise to the bare region around the isthmus of the pharynx. (See Figure 2 in color.)
FIGURE 3. View of the ventral ganglion. The cells of the ventral ganglion are bounded by a basal lamina, which separates them from cells of the lateral ganglia even though they are adjacent (figure 2). The posterior region of the ganglion is interrupted by the presence of the excretory duct and excretory canal cells, which exclude the cell bodies of neurons from this region. VB2, AVFR and SABVL are part of the retro-vesicular ganglion and are separated from the cells of the ventral ganglion by a basal lamina. All the cells of the ventral ganglion project into the nerve ring, and several of the cell classes present also have members in the lateral ganglia.
Most of the neurons of C. elegans have their cell bodies situated in the head around the pharynx (figure 2). The pharynx is composed of two prominent bulbs joined by an isthmus. An extensive region of neuropile, the circumpharyngeal nerve ring, encircles the centre region of the isthmus and has cell bodies clustered adjacent to it both anteriorly and posteriorly. There are no obvious sub groupings of the neuron cell bodies anterior to the ring and so these have been lumped together and referred to as the anterior ganglion. The anterior ganglion is mainly made up of the cell bodies of neurons, sheath cells and socket cells from the sensilla that are located in the six labia of the head (figure 1). The relative positions of cell bodies within ganglia are fairly well conserved between animals of the same developmental stage and genotype. There is a certain amount of `slop' however; the extent of this can be seen by comparing the left and right sides illustrated in figure 2. The most extreme cases of variability in this region arise because the anterior bulb of the pharynx fits fairly tightly in the body cavity and excludes cell bodies from its region of maximum diameter. This leads to some uncertainty in the position of some cell bodies with respect to the bulb; for example, in the N2U reconstruction, OLQsoDL lies anterior to the bulb, whereas its symmetrical partner, OLQsoDR, lies posterior to the bulb (figure 2). In live animals, cells can sometimes be seen to flip from one side of the anterior bulb to the other as the pharynx moves.
Posterior to the nerve ring, the basal laminae split the cell bodies adjacent to the ring into four groups (figure 13): a small dorsal ganglion, two lateral ganglia, and a ventral ganglion (figure 3). All receptor neurons of the amphid sensilla have their cell bodies in the lateral ganglia, which also contain cell bodies of motoneurons and interneurons. The dorsal ganglion contains interneurons together with the neurons of the two dorsal cephalic sensilla. The ventral ganglion contains interneurons and motoneurons. The cell bodies of the ventral ganglion are separated into two groups (figure 3) by a mechanical intrusion, as are the cells of the anterior ganglion. In this case it is the excretory duct and canal that displaces the cells.
The posterior extremities of the ventral ganglion overlap the anterior of the retrovesicular ganglion, which is situated on the ventral mid-line posterior to the excretory pore (figure 2); however, the two groups of cells are distinct, being separated by basal laminae. A single row of cell bodies runs down the ventral mid-line (figure 4) from the retro-vesicular ganglion to the tail, where it ends in another ganglion, the pre-anal ganglion. There are three extra ganglia in the tail: two laterally symmetric lumbar ganglia and a single, small dorso-rectal ganglion (figure 5). There is a pair of small lateral ganglia in the posterior body, the posterior lateral ganglia, and there are some isolated cells along the body laterally (figure 4).
FIGURE 4. The locations of the cell bodies of all the neurons and their associated cells in the body are shown on the left-hand side (a), the right-hand side (b) and the middle (c). These diagrams were derived from light microscope observations (Sulston & Horvitz 1977). The asymmetrics in the positions of SDQL/R, AVM and PVM are a consequence of the different migration patterns of the initially bilaterally symmetric precursor cells QL and QR. The ventral cord motoneurons shown in (c) can be separated into those that are present at hatching, shown by the labels on the right, and those that develop postembryonically, shown by the labels on the left. Thc anterior-posterior sequence of cell types in these two groups is always the same, but there is some slight variation in the way the two groups intercalate, giving some variation in the combined adult sequence.
The anterior ganglion, the ventral ganglion and the dorso-rectal ganglion are completely bounded; that is, they have clear structurally defined limits to their extents. The others are 'open' in that there are no specific boundaries at one end. The retrovesicular ganglion is open and continuous with the region containing the motoneurons of the ventral cord, which in turn is open and continuous with the pre-anal ganglion. Similarly, the lateral ganglia are open and continuous with the isolated cells on the lateral lines, the posterior lateral ganglia and the lumbar ganglia. Thus the body has three main compartments where neuron cell bodies are located, two lateral and one ventral.
There seem to be no functional correlates to the groupings of cells into particular ganglia. Often cells are more analogous, in structure and connectivity, to cells in other ganglia than to cells in the same ganglia. Ganglia simply seem to be local groupings of cell bodies brought about by extraneous mechanical factors.
Disposition of process tracts The nervous system of C. elegans is made up of a set of interconnected parallel process bundles. These run either longitudinally or circumferentially, adjacent to hypodermal tissue (figures 6 and 7). The two sub-dorsal and the two sub-ventral labia at the tip of the head each have a single process bundle associated with them. This is made up of processes from the sensilla in the labium, together with other processes that terminate near the sensilla but have no differentiated endings. The lateral labia have similar process bundles but, in addition, each has a larger process bundle made up of processes of the neurons of the amphid sensilla. Most of the processes in the six non-amphidial bundles have associated cell bodies, which are situated in front of the nerve ring in the anterior ganglion. Individual processes peel away from the bundle to join their (bipolar) cell bodies. A second, posteriorly directed process emanates from the cell body and rejoins the process bundle, running in the same region of the bundle as its anteriorly directed counterpart. The six labial process bundles run posteriorly past the outside of the nerve ring and then turn to enter the nerve ring near its posterior face (figure 6). The processes in the amphid bundle bypass the ring completely and run to their (bipolar) cell bodies, situated in the lateral ganglia. Axonal processes from these cell bodies, along with processes from monopolar cell bodies of interneurons and motoneurons, enter the nerve ring via two main routes. Cells in the ventral region of the lateral ganglia have processes that join the amphidial commissures; these run circumferentially round the animal, between muscle and hypodermis, to the ventral mid-line, where they turn and enter the nerve ring. Cells in the dorsal regions of the lateral ganglia do not take this somewhat circuitous route but enter the nerve ring directly sub-dorsally.
FIGURE 6. Process tracts in the head. A process from a neuron generally runs in a bundle along with the processes of other neurons. These process bundles either run longitudinally or as commissures circumferentially. The six labia at the tip of the head are richly endowed with sensory receptors. Processes from these receptors, along with those from some additional neurons, run along the labial process bundles to their cell bodies, which are situated anteriorly to the nerve ring. Posteriorly directed processes from these bipolar cell bodies rejoin the process bundles and pass along the outside surface of the nerve ring. They then turn and enter the posterior regions of the ring neuropile and move to the inside surface of the ring, and, turning again, they run to the anterior regions of the ring neuropile, where they disperse and have most of their synaptic interactions. Each lateral labium also has an amphid sensillum, and process bundles from its component neurons run posteriorly past the nerve ring to their bipolar cell bodies in the lateral ganglia. Most of these neurons send processes into the ventral cord via the amphid commissures, which then project into the nerve ring. The deirid receptors, along with several other neurons with cell bodies in the posterior regions of the lateral ganglia, send processes into the ventral cord via the deirid commissures. The ventral cord (figure 18) is the main process bundle that emanates from the nerve ring and contains processes of interneurons and motoneurons. Most of the processes in the dorsal cord originate in the ventral cord and enter the dorsal cord via commissures. There are four sub-lateral process bundles, made up of processes from motoneurons and interneurons that come from the nerve ring. These run anteriorly and posteriorly from the nerve ring and eventually end (figure 8). (See Figure 6 in color.)
FIGURE 7. Left-hand (a) and right-hand (b) process tracts in the body. The main process tracts are the ventral cord, the dorsal cord, the excretory canal associated processes and the posteriorly directed sub-lateral processes. The ventral cord consists of processes of interneurons and processes and cell bodies of motoneurons (figures 4 and 18). The ventral cord bifurcates at the anus and runs up to the lumbar ganglia via the lumbar commissures. The dorsal cord (figure 19) is predominantly made up of motoneuron processes that have come from the ventral cord via circumferential commissures, which are distributed along the length of the body. Most of the processes in the posterior sub-lateral cords are derived from the nerve ring. These process bundles run sub-laterally under the body muscles (figure 8) anteriorly, but move laterally to each side of the lateral hypodermal ridges where most of the processes end. Processes from SDQ and PLN run into these cords from the opposite direction from laterally situated cell bodies. The processes of CAN, ALA, PVD and also (in the anterior of the animal) BDU, run together alongside the excretory cell for most of its length. The anterior touch receptors, ALM, together with their associated neurons, ALN, run anteriorly near the dorsal side of the lateral hypodermal ridges; their posterior counterparts, PLM and PLN, run anteriorly near the ventral side of the ridges.
In the anterior body there are four sub-laterally situated process bundles that run underneath the body muscles (figure 6). They run in a straight line approximately corresponding to the junction of the two rows of muscles in each quadrant. There are five processes in the sub-lateral cords behind the nerve ring, and two in each of the cords in front of the ring (figure 8). The processes in the anterior cords peter out in the head; those in the posterior cord move to a more lateral position near the middle of the body, where most of them end (figure 7).
Three nerve processes run for much of the length of the animal, closely associated with the excretory canal (CAN-a). These processes run into the nerve ring at the anterior end (figure 6) and peter out posteriorly in the tail.
The only remaining process bundles in the body are those made by the lateral touch receptor neurons, ALM and PLM, and their associated neurons, ALN and PLN. The anterior touch receptors, ALM, run in close association with ALN near the dorsal margin of the lateral hypodermal ridges, whereas the posterior receptors, PLM, run along with PLN near the ventral margin of the lateral hypodermal ridges. The processes of ALN maintain their dorso-lateral location in the posterior part of the body although they are not in close association with ALM in this region.
FIGURE 8. Most of the sub-lateral processes originate from the nerve ring and run longitudinally underneath the muscle quadrants close to the line of apposition of the two muscle rows. Apart from a single NMJ, no synapses have been seen on these processes. There are two processes in each of the sub-lateral cords anterior to the ring (a) and five in each of the cords posterior to the ring (b). The individual processes run in fixed positions within the cords. The posterior cords include processes from PLN and SDQ, which must have grown in the opposite direction to the others, as their cell bodies are situated laterally in the body (figure 7). Apart from the processes of the cells, the sublateral processes eventually peter out (figure 6 and 7).
Musculature Nematode body muscles are unusual in that their sarcomeres have an oblique conformation with the actomyosin filaments, aligned at an angle of about C 10 to the Z lines, rather than being orthogonal to them. This type of arrangement has been referred to as obliquely striated muscle (Rosenbluth 1965; Waterston et al. 1980). The Z lines consist of longitudinally oriented lines of discrete structures (dense bodies), which are darkly staining in electron micrographs (figure 18). These structures are roughly conical in shape; the base of the cone is adjacent to the cell membrane, which is in turn adjacent to the hypodermis and cuticle. The body muscles probably have attachments to the elastic cuticle distributed along their length, since no specialized focal attachment points are seen at the end of these muscle cells.
Body muscles are rhomboid-shaped and are arranged as two parallel rows in each quadrant (figure 10). There are 95 muscle cells in the adult; the left ventral quadrant contains 23 and the other quadrants each contain 24 (Sulston & Horvitz 1977). The muscles in the body can be divided up into three groups on the basis of their source of synaptic input: the anterior group of four muscles in each quadrant, innervated by motoneurons in the nerve ring, the next group of four, which is dually innervated by motoneurons in the nerve ring and ventral cord, and the remaining muscles, which are innervated solely by the motoneurons of the ventral cord (figure 10; see also Ware et al. 1975).
Motoneurons of the ventral cord innervate either both dorsal or both ventral quadrants of muscle. The body can therefore only propagate dorso-ventral waves during locomotion. The head, on the other hand, can make lateral as well as dorso-ventral movements when the animal is foraging. This is probably because the motoneurons in the nerve ring do not synapse onto two quadrants of muscles, but instead are restricted to two adjacent rows (not necessarily in the same quadrant). This would allow differential activation of muscles in adjacent quadrants and possibly even in adjacent rows.
Nematode muscles are unusual in that they have neuron-like processes that run from the muscle bellies to the neuron process bundles in which motoneuron axons reside (figures 9 and 18). Neuromuscular junctions (NMJs) are made by axons running along the surface of their process bundle, through the bounding basal lamina of the bundle and onto muscle arms (see, for example, VDn-a). Muscle arms interdigitate extensively and crowd round regions where NMJs occur; there are often gap junctions between the arms in these regions. Muscle arms in the body converge at the dorsal and ventral mid-lines, where they interdigitate and contact the dorsal and ventral cords (figure 9). Arms from the head muscles, which receive their innervation from motoneurons in the nerve ring, run down past the outside of the ring and then turn and run anteriorly, closely apposed to the inner surface of the ring. Here they sort out in such a way that arms from each muscle row make an arc of about 45 degrees (figure 15). Thus there is a mapping by the muscle arms of the spatial organization of the muscle cells onto the inner surface of the nerve ring. Motoneuron axons run adjacent to the inside surface of the ring and are arranged in a well-ordered pattern (figure 14). The inside surface of the muscle- arm complex in the region of the NMJs is lined by the thin sheet-like processes of the GLR cells (figures 14 and 15). No chemical synapses are seen on these cells, so they are probably not neuronal; however, they do make gap junctions to muscle arms and to RME motoneurons (figure 15).
FIGURE 9. The body consists of a tube of hypodermal tissue made up of two cell types: the syncytial cell (SY), which makes up the dorsal and ventral hypodermis, and the lateral seam cells (SE), which are also syncytial in the adult and are joined to the syncytial cell by desmosomes. Longitudinal ridges of hypodermis run down the body on the lateral, dorsal and ventral lines. Process bundles that make up the dorsal and ventral cords run alongside the dorsal and ventral hypodermal ridges and are separated from the pseudocoelome by a basal lamina. Thc body musculature consists of four quadrants of obliquely striated muscles. Each quadrant consists of two closely apposed rows of muscle cells. The motoneurons that innervate body muscles have longitudinal unbranched processes which are confined to the dorsal and ventral cords. Muscle cells send out processes to the nerve cords, where motoneurons synapse onto them through the basal lamina at NMJs.
FIGURE 10. 'Orange peel' projection of muscles in the head. The reconstruction was derived from electron micrographs. The muscles are organized as longitudinal strips in each of the four body quadrants (figure 9). Each quadrant has two adjacent rows of muscle cells. The muscles are obliquely striated and packed diagonally so that the sarcomeres are oriented longitudinally. The first two muscle cells in the two ventral and two dorsal rows are smaller than their lateral counterparts, giving a stagger to the packing of the two rows of cells in a quadrant. The first four muscles in each quadrant are innervated exclusively by motoneurons in the nerve ring. The second block of four muscles is dually innervated, receiving synaptic input from motoneurons in the nerve ring and the anterior ventral cord. The rest of the muscles in the body are exclusively innervated by NMJs in the dorsal and ventral cords (figure 9). The eight muscle rows have been labelled dorso-medial right (DMR), dorso-lateral right (DLR), ventro-lateral right (VLR), ventro-medial right (VMR), ventro-medial left (VML), ventro-lateral left (VLL), dorso-lateral left (DLL) and dorso-medial left (DML).
There are sixteen sex-specific muscles in the hermaphrodite; eight are associated with the uterus and eight with the vulva (figure 11). Unlike the body muscles, these muscles have focal attachment points at their ends and do not have obliquely oriented sarcomeres. The hermaphrodite gonad has twofold rotational symmetry, the axis of symmetry passing through the centre of the vulva. The uterine muscles distal to the vulva, um2, wrap round the uterus, whereas the uterine muscles proximal to the vulva, um1, attach to the lateral lines. Both sets of muscles consist of a pair of muscles that are joined at the ventral mid-line. There are two sets of four vulval muscles, vm1 and vm2. The vm1 muscles are attached to the body wall sub-ventrally, insinuating themselves between the rows of body muscles, and are attached at their proximal ends to the hypodermal lips of the vulva. The vm2 muscles attach to the body more ventrally, at the ventral margin of the muscle quadrants, and are attached at their proximal ends to the opening in the uterus, which connects to the vulva. Most of the synaptic input to the vulval muscles comes from VCn and HSN neurons and is directed onto the vm2 muscles (figure 11 c ). The other muscles are either directly or indirectly connected to vm2 via gap junctions. The vm1R muscles send a muscle arm down to the ventral cord, where it receives a small amount of synaptic input from ventral cord motoneurons.
Defecation is controlled by three sets of muscles: the anal depressor muscle, the sphincter muscle and two laterally symmetric intestinal muscles (figure 12). The anal depressor muscle is a large H-shaped muscle, which lifts the roof of the anus when it contracts. The sphincter muscle is a circular muscle that closes off the end of the gut. The intestinal muscles have longitudinally oriented filaments, which are situated in the ventral regions of the cells. The dorsal regions flatten into thin sheets, which wrap round the posterior ventral regions of the intestine and are probably attached to it. Muscle arms from these three sets of muscles run into the pre-anal ganglion and are coupled together via gap junctions. Surprisingly little synaptic input was found to be present on the defecation muscles, with only a single NMJ being made by DVB.
FIGURE 11. Egg laying is controlled by a set of sixteen muscle cells in the hermaphrodite, eight of which act to squeeze the uterus (a) and eight to open the vulva (b). The distal uterine muscles, um2, form circumferential bands of muscle round the distal regions of the uterus. The um1 muscles attach to the lateral hypodermis and wrap round the proximal ventral regions of the uterus. The vm1 muscles attach to the body hypodermis at the ventro-lateral body muscle margins and at the vulval opening. The vm2 muscles attach to the body hypodermis sub-laterally, insinuating themselves between the body muscles, and to the uterus at the vulval opening. The vulval and uterine muscles have gap junctions to each other, as shown in (c). The main synaptic input is onto the vm2 muscles and comes from VCn (*a) and HSN (*a). The NMJs are dorsal to the main part of the ventral cord (VCn-a). vm1R sends an arm down into the ventral cord and receives single synapses from VD7, VB6. and VA7.
FIGURE 12. There are three muscles directly involved in defecation: the anal depressor muscle, the anal sphincter muscle and the two intestinal muscles. The anal depressor muscle is a large H-shaped cell, which lifts the posterior dorsal surface of the rectum so as to open it and discharge its contents. The intestinal muscles have longitudinally oriented contractile filaments and attach to the body hypodermis at the ventral muscle margin and to the intestine via several distributed contacts on its ventral surface. The intestinal and depressor muscles send muscle arms to the posterior regions of the pre-anal ganglion, where they receive synaptic input from DVB (*c).
Basal lamina The pseudocoelomic cavity is lined with a thin (20 nm) basal lamina, which effectively separates the muscles from the hypodermal and nervous tissues. This lamina has an anisotropic structure, as parallel striations with a spacing of 30 nm can be seen when it is sectioned obliquely (White et al. 1976). The gonad and the gut are ensheathed by similar basal laminae; the pharynx is ensheathed by its own, rather thicker (45 nm) basal lamina (Albertson & Thomson 1976). The dorsal and ventral nerve cords, together with their respective hypodermal ridges, are bounded by the pseudocoelomic basal lamina (figures 18 and 19); the lateral hypodermal ridges and the laterally located ganglia are similarly bounded. The boundary curves smoothly, suggesting that the lamina may be under tension in these regions.
All the nervous system is situated to one side of the pseudocoelomic basal lamina, with the exception of the cell bodies of URX, CEPD and GLR. The processes of URX and CEPD run together on each side as they leave the ring sub-dorsally. They are surrounded by, and eventually penetrate, the basal lamina in these regions before reaching their cell bodies, which are situated in the pseudocoelomic cavity. The basal lamina may also be penetrated in four places on the inside of the nerve ring by muscle arms (figure 14 and RIM-d). This enables a motoneuron (RIM), which has its axon buried in the interior of the ring neuropile, to make NMJs.
Nerve processes seem to be constrained to run alongside the lamina. Processes that run from the ventral to the dorsal cord, for example, run round the animal, travelling underneath the muscle quadrants instead of taking a more direct internal route. In the main part of the body cavity the dorsal and ventral ridges are quite small, consisting of a ridge of hypodermis and an adjacent process bundle (figure 13d). As the head is approached, the dorsal, ventral and lateral ridges enlarge as they become filled with the cell bodies of their respective ganglia (figure 13c). Eventually the basal laminae bounding the four ridges meet and fuse (figure 13b). An internal tract is now opened up and processes course round it inside the muscle quadrants forming the nerve ring. This organization is maintained up to the tip of the head with the four muscle quadrants running in tubes of basal laminae (figure 13a). The central ring of lamina left after the ridges have fused ends in the vicinity of the nerve ring. It appears to terminate on the cylinder that is made up of the sheet-like processes of the GLR cells. This structure is situated on the inside of the nerve ring between the pharynx and the muscle arms.
The arrangement of the basal lamina lining the pseudocoelome suggests that it may be instrumental in the establishment of the general topography of process tracts in the nervous system. Processes from neurons have been shown to grow preferentially along ordered fibrillar arrays (Weiss 1934). The striated structure may likewise serve to guide initial process outgrowths, thereby establishing the antero-posterior and circumferential system of process bundles that are a feature of the nervous system of C. elegans.
FIGURE 13. The pseudocoelome in the body is bounded by a basal lamina, which covers all the hypodermal and nervous tissue (d). The muscles are in the pseudocoelomic cavity. Processes of neurons do not, in general, cross the basal lamina. Commissures between the dorsal and ventral cords pass underneath the muscle quadrants and do not enter the pseudocoelomic cavity. As thc ring is approached, the dorsal, ventral and lateral cords enlarge where they are filled with cell bodies of the respective ganglia (c). There is no direct route between the ganglia at this point, however, and cell bodies in the lateral ganglia send processes into the ventral cord via the amphidial commissures (figure 6). At the level of the nerve ring, the lobes of the basal lamina fuse inside the muscle quadrants (b) allowing thc processes in the nerve ring to run round without having to pass underneath the muscle quadrants. The processes of the nerve ring, like those of the nerve cords, run along side a ridge of hypodermis (a), which is anterior to the neuropile. The nerve ring seals off the anterior end of the pseudocoelomic cavity and there is no basal lamina bounding the hypodermal and nervous tissue in the head, except for that bounding the pharynx.
Neurons
Branching structure
The component neurons of the nervous system of C. elegans have simple, unbranched morphologies. Few neurons have more than two processes, and many are monopolar with only a single process (see, for example, AIA). Processes of neurons run in parallel bundles except in the immediate vicinity of their cell bodies, where they join the bundle. This region is not extensive, however, as cell bodies are generally situated close to the bundle into which they project. Branching typically occurs when a neuron has a process that leaves the main bundle to run out as a commissure (see, for example, VDn), or at a discontinuity, where one bundle joins another (as in AQR where it leaves the ventral cord and enters the nerve ring).
Neurons with a branched structure generally have very similar patterns of branching in different animals; however, there are a few interesting cases where differences occur between animals, or between sides of the same animal. The interneuron RID lies on the dorsal mid-line and sends a process round the left-hand side of the nerve ring in the N2U animal and round the right-hand side in the JSH animal. The nerve ring has a high degree of bilateral symmetry and the process of RID runs in a similar position relative to the neighbouring processes whether it runs on the left or the right.
The interneuron PVN is the most highly branched class of neuron in C. elegans. The main processes of PVN run up the ventral cord and enter the nerve ring on the right-hand side, travelling round it in an anticlockwise direction. PVNL has an additional branch, which separates from the main process at a point behind the excretory duct. This branch enters the ring on the left-hand side, travelling round it in a clockwise direction. This process (which is not present on PVNR) runs in the same region of neuropile as do the main processes of both PVNR and PVNL, which are travelling in the opposite direction; they also make similar synaptic contacts. Other examples of such conservative variation in branching patterns have previously been noted in the cephalic receptor neurons, CEP (Sulston et al. 1975). These observations suggest that, irrespective of branching structure or even direction of growth, a process is capable of locating its appropriate neighbourhood within the neuropile and forming its characteristic synaptic connections.
A few examples of non-conservative changes in branching pattern have been seen. A fairly major branch is missing on RMFR in the N2U animal but is present on its contralateral partner and is also present on RMFR in the JSH animal. As the missing process has all the NMJs made by this motoneuron, such a change must have a profound effect on the function of RMFR in this instance. It seems reasonable to consider such incidences of branching failures as developmental errors in the construction of the nervous system, which could perhaps give rise to non-genetically related variations in behaviour between animals.
Branch termination
The processes of many classes of neuron terminate at the point of contact with a process from a neighbouring member of the same class. There is usually a gap junction at this point (as in ASI on the dorsal mid-line), although there is one case where processes touch and terminate with no gap junction (RIF). There are also a few cases where such contact terminations can occur between heterologous classes (e.g. between processes of ALM and AVM in the nerve ring). The most striking examples of contact termination are exhibited by the DDn and the VDn motoneurons of the ventral cord. There are six DDns and thirteen VDns evenly distributed along the length of the cord. Each of these classes has processes in both ventral and dorsal cords. Together, their processes make an unbroken line of non-overlapping processes in each cord (White et al. 1976). This behaviour seems to be an intrinsic property of certain classes of neuron; other classes of neuron make contacts and gap junctions with members of their own class but do not terminate at the site of initial contact and may have considerable overlap (see, for example, ASE, AIN).
Gap junctions
Gap junctions are organelles that mediate electrical and metabolic coupling between cells (Bennett 1977). They are seen in C. elegans as regions where the membranes from two adjacent cells are closely apposed and appear more darkly staining than surrounding regions (as in VBn-c). When gap junctions are sectioned transversely, a gap of about 8 nm can be seen separating the membranes. The region of close apposition is usually in the form of a plaque of about 350 nm diameter. The membranes at the junction are notably flatter than those of the surrounding regions. The gap junctions seen in C. elegans resemble those described by Pappas & Waxman (1972).
Gap junctions are seen between muscle cells and between neurons. Apart from a couple of possible exceptions (RMD-h and VCn-f), gap junctions are not seen between muscle cells and neurons, probably because there is usually a basal lamina separating the two. The glial-like cells, GLR, are unique in that they make gap junctions to both muscles (GLR-c) and neurons (GLR-d). They do not, however, make gap junctions to themselves. The arrangement of these gap junctions is shown in figure 15.
Muscle arms from muscles in the head have a striking arrangement of gap junctions where they interdigitate at the inside of the nerve ring. Arms make gap junctions with arms from muscle cells in adjacent quadrants but not with arms from muscle cells in the same quadrant, even though both sets of arms are equally accessible (figure 15). Muscles in the same quadrant are, however, connected by gap junctions, but the connections are situated in the region of the muscle cell bellies, well away from the arms. Thus it seems as though muscle arms, when they grow into the nerve ring, can discriminate between the arms of muscle cells that are already connected to themselves via gap junctions and those that are not.
Chemical synapses
Chemical synapses in C. elegans occur en passant between neighbouring parallel processes. The presynaptic process has a vesicle-filled varicosity and a specialized, darkly staining region in the membrane adjacent to the point of contact with the postsynaptic elements (see, for example, EAG-a). A considerable variation in the size of the presynaptic regions was found (compare OLQ-a with PVN-a). The presynaptic specializations also vary in prominence between different classes of synaptic contact in a way that does not necessarily correspond to the size or the number of vesicles in the presynaptic process. The extremes of this variation are represented by RIP, on the one hand, which has structures that look like presynaptic specializations but with no associated synaptic vesicles (RIP-a); and, on the other, by DVA, which has large vesicle-filled varicosities but rather small presynaptic specializations (DVA-b). There is also considerable variation in the number of chemical synapses between pairs of interacting processes. There are many cases where there is only a single synapse present. At the other end of the scale, the largest number of synapses seen between processes is nineteen (AVDL onto AVAR); more typically it is around five. Some of the single synapses that are seen are small, with few synaptic vesicles or indistinct presynaptic specializations. Synapses of this type are also rather variable, in that they are not present in some individuals and therefore probably not very significant. On the other hand, some single synapses are large, with many vesicles and unambiguous presynaptic specializations. These synapses are seen in all individuals and so are probably significant. This latter type of synapse seems to occur when the layout of the two interacting processes is such that they are only adjacent for a limited extent. In these cases there may only be room for a single synapse in the region where the two processes are adjacent.
Although the fixation and staining procedures that were used are not optimal- for the preservation and visualization of vesicle morphology, several classes of vesicle can be clearly distinguished. The most ubiquitous vesicles are spherical, 35 nm in diameter, and have lightly staining interiors (see, for example, RIA-a). Some classes of neuron, including most of the amphid receptors, have a second class of vesicle coexisting with vesicles of this first type. These vesicles are larger and have darkly staining cores (as in ASK-a); the relative proportions of the two types of vesicle varies with cell class. There is a certain amount of variation in the staining properties of these dark-cored vesicles between classes; the sizes also vary, ranging from 37 nm (ASE) to 53 nm (ASK). The dark-cored vesicles seem generally to be excluded from the region immediately adjacent to the presynaptic specialization, which contains only the smaller type of vesicle. A similar segregation of vesicle types is exhibited by DVA, which has a large process in the nerve ring, filled with irregularly shaped vesicles, but has small spherical vesicles next to presynaptic specializations (DVA-a). The neurotransmitters that may be contained in the dark-cored vesicles are not known. Dopamine has been shown to be present in CEP, ADE and PDE neurons (Sulston et al. 1975). Acetylcholine is probably used as a neurotransmitter by the ventral cord motoneurons VAn, VEn, DAn, DEn and ASn, as this transmitter has been shown to be used in the equivalent neurons in Ascaris (Johnson & Stretton 1980). All these classes of neuron have uniform populations of spherical, 35 nm, synaptic vesicles, with no dark-cored vesicles present (see, for example, CEP-a, VAn-a).
Chemical synapses in C. elegans usually have no visible specializations on postsynaptic elements and consequently there is often some ambiguity as to the identities of these elements. In some cases, the disposition of the processes is such that there clearly can be only one postsynaptic element (as in ASE-a). In many other cases there are two (for example, in ADF-a) or, more rarely, three (for example, in AIY-e) postsynaptic elements, making a dyadic or triadic synapse (Dowling & Boycott 1966). It was difficult to know in these cases whether all the postsynaptic elements are functional (i.e. have an appropriate receptor) or are just neighbouring processes. It seems likely that, in many cases, all the possible postsynaptic elements could be functional, as particular dyadic or triadic combinations are found to occur in many instances (for example, AIA and AIB are often the two postsynaptic elements in a dyadic synapse). Some synaptic pairings are only seen in the context of multiple synapses. Although this may suggest that such a pairing could be non-functional, there are cases where this cannot be so, as the other postsynaptic element of the dyadic synapse is also seen only in the context of a multiple synapse (for example, RIB and AVE are postsynaptic to AUA, and AVE and AIZ are postsynaptic to RIG). This observation raises the interesting possibility that, in some cases, synaptogenesis may be dependent on the simultaneous presence of two particular postsynaptic elements.
Several process pairs are seen to synapse onto each other reciprocally. AVAL/R and PVCL/R synapse onto each other along the length of the ventral cord, for example, but there is no particular spatial relation between the two types of synapse. The reciprocal synapses made by RIA and RMD are usually situated close to each other, however, making a characteristic structure (RIA-e). Such an organization may provide positive or negative feedback in these synaptic connections.
Many classes of neuron are found to have regions of process that are devoid of presynaptic specializations. This could be because the particular class of neuron does not have many synapses in total or that these regions corresponded to regions where there are no suitable postsynaptic partners. In several cases neither of these explanations can be valid. The interneurons AVA, AVB, AVD and AVE are all exclusively postsynaptic in the nerve ring, yet they have extensive synaptic outputs in the ventral cord. Furthermore, AVD, AVE and AVB all have extensive synapses onto AVA along the cord; however, in the nerve ring, processes from these cells do not make such synapses even though they are accessible to AVA (i.e. are adjacent to its processes) for part of their extent within the ring. Thus it appears that certain classes of neuron can localize the regions where they are presynaptic. Those regions of process that are devoid of presynaptic contacts are often more lightly stained than adjacent processes (AVA-a). There seems to be no localization of postsynaptic contacts.
Occasionally, presynaptic elements are seen with no obvious postsynaptic partner, or with a hypodermal cell as the only possible partner. AVB is particularly prone to this behaviour, having six such structures along the length of the ventral cord (see, for example, AVB-a). It is difficult to know how to interpret these structures; they could possibly be functional synapses and control some hypodermal cell function such as cuticle deposition or moulting, or they could be artefacts.
Neuromuscular junctions
Neuromuscular junctions (NMJs) are special cases of chemical synapses where at least one of the postsynaptic elements is muscle. As the muscle and nervous system are situated on opposite sides of the pseudocoelomic basal lamina, NMJs have to pass through the lamina with the presynaptic elements (the motoneuron axons) on one side and the main postsynaptic elements (the muscle arms) on the other. Because of this arrangement, NMJs are constrained to lie on the two-dimensional surface of the lamina. NMJs usually have several postsynaptic elements. On the inside of the nerve ring, there is a continuous plexus of arms from muscles in the head and a high density of NMJs (figure 14). In the ventral cord, the NMJs are more dispersed and muscle arms crowd round and interdigitate at foci where there are presynaptic elaborations on motoneuron axons (figure 18).
Certain classes of neuron (VDn, DDn, RMD, SMD, RME and RIP) have processes that are postsynaptic at NMJs. These processes are on the same side of the basal lamina as the presynaptic elements and often have a short branch, which dips in and intercepts the NMJ (see, for example, RMD-a). Because of this behaviour, it seems likely that these processes are functional postsynaptic elements. The disposition of the dendritic processes relative to the NMJs that they are intercepting suggests that the NMJs might have formed first and the dendrites might have moved in and insinuated themselves into position later. There is likely to be some specificity as to which NMJs are intercepted by particular dendrites, as dendrites along the ventral cord are not associated with the NMJs of VDn and DDn, but are associated with the NMJs of the other motoneuron classes active in the nerve cord, even though all classes of NMJ are equally accessible to the dendritic processes.
With the exception of RIP, all the classes of neuron that have postsynaptic elements in NMJs are motoneurons themselves and, interestingly, have NMJs on the diametrically opposite side of the animal to the regions where they are postsynaptic. Thus it seems likely that these classes of neuron act as cross-inhibitors, ensuring that muscle contractions in diametrically opposite regions of the animal operate in antiphase. Neurons analogous to VDn and DDn have been identified in the ventral cord of A. lumbricoides and have been shown to be inhibitory (Johnson & Stretton 1980).
The arrangement of motoneuron axons around the inside surface of the nerve ring was found to be the most highly ordered region of neuropile in the nervous system (figure 14). The ordering is such that it is often possible to identify many of the processes in this region by their appearance in a single appropriately positioned section. Several of the NMJ sin this region are organized as characteristic complexes made up of presynaptic endings clustered around a dendritic process (figure 14). The dendritic processes are from RMD, SMD and RIP. The NMJs made by RMD and SMD are situated diametrically opposite their dendritic processes. The RIP neurons also have processes that cross over to the diametrically opposite side from the dendritic regions, even though they are not motoneurons. These processes eventually enter the pharynx (Ward et al. 1975; Albertson & Thomson 1976).
The arms from each row of head muscles are arranged around the inside surface of the nerve ring such that arms from each row occupy a well-defined arc. This arc is positioned in an equivalent location to that of the muscle row from which the arms originated (figure 15). There is thus a fairly precise mapping of the circumferential positions of the muscle rows, by the muscle arms, onto the motor endplate region. The ordering of the motoneuron axons on one side of the basal lamina and the muscle arms on the other is highest at the regions immediately adjacent to the lamina but is less apparent away from it.
The flattened processes of the GLR cells cover the inside surface of the plexus of muscle arms inside the nerve ring and are seen to have gap junctions with adjacent muscle arms (figure 15). The processes of GLR are found to be aligned with the arcs of muscle arms from each row (figure 14). The sub-dorsal and sub-ventral sets (GLRDL/R and GLRVL/R) are each associated with muscle arms from one row, whereas the lateral pair (GLRL/R) are larger in circumference and are each associated with two muscle rows. The points of contact between adjacent GLR processes are closely aligned with the points of contact of the arcs of muscle arms, except in the case of the muscle rows lying either side of the lateral lines. In these, there is no GLR process junction and a certain amount of mixing of the muscle arms at the point of contact of adjacent arcs occurs, whereas there is no mixing at the points of contact that have an associated GLR process junction. These observations suggest that the GLR processes may act to guide muscle arms and confine them to their appropriate territories on the inside of the nerve ending.
FIGURE 14. Neuromuscular junctions in the nerve ring. The eight rows of muscles in the head and neck (figure 10) have muscle arms that project onto the inside surface of the nerve ring in a highly ordered way (figure 15). They are sandwiched between the thin sheet-like processes of GLR cells on the inside and the motoneurons of the nerve ring on the outside. Four spurs of muscle arm penetrate into the anterior neuropile of the ring sub-laterally and receive synaptic inputs from RIM, which runs in the interior of the ring neuropile. The other classes of motoneuron form complex, but well-defined, structures adjacent to the inner surface of the nerve ring. Most NMJs are dyadic, with dendrites of other motoneuron classes or RIP as the corecipients. The dots in the processes show the locations of the presynaptic specializations.
FIGURE 15. Head and neck muscle projections. The muscle arms from thc 32 head and neck muscles send arm posteriorly past the outside surface of the nerve ring. These then turn and run anteriorly onto the inside surface of the ring. The muscle arms are highly ordered in this region and map onto the inside surface according to the circumferential location of the muscle bellies. Muscle arms have gap junctions to arms from adjacent muscles in neighbouring quadrants and to GLR cells. RME motoneurons also have gap junctions to GLR cells in thc arrangement shown. There arc gap junctions between the muscle bellies of muscles in adjacent rows of the same quadrant but, interestingly, none are seen between the arms from these muscles, even though they interdigitate extensively.
One motoneuron, RIM, is unusual in that it does not have its axon adjacent to the inside surface of the nerve ring. Instead it forms NMJs onto four spurs of muscle arms that invade the neuropile of the ring (figure 14). It is difficult to visualize the basal lamina in these regions, so it is not clear whether the muscle arms actually penetrate the basal lamina at these points or whether the basal lamina is herniated. The sites of these invaginations again correspond to junctions between GLR processes and are fairly small; muscle arms anterior and posterior to these regions run along the inside surface of the ring.
There are seven main classes of motoneuron in the ventral cord: VAn, DAn, VEn, DEn, ASn, VDn and DDn. Members of each class are evenly distributed along the length of the cord (White et at. 1976). Within each class there are sharply defined transition points where one axon becomes synaptically active, having many NMJs along the cord, and the adjacent axon becomes inactive, having no more NMJs. These transition points occur in slightly different positions for each class; such observations suggest that there might be intraclass competition for territory along the ventral cord (White et al. 1976). Similar intraclass competitions for territories have been shown to occur in two dimensions for classes of ganglion cell in the vertebrate retina (Wassle et al. 1981). In the nerve ring, many of the motoneuron classes have NMJs at discrete points around the motor end plate region and so it seems unlikely that intraclass competition has a role in establishing NMJ territories in these cases. The RMDD/V motoneurons have NMJs around the whole circumference of the ring, however, with abrupt transitions between adjacent class members, which each have NMJs over a 45° arc. Thus it seems possible that, in this case, intraclass competition may be used to partition out territory for NMJs to the class members.
The organization of processes within bundles
The process bundles in C. elegans are spatially ordered, with processes running in characteristic positions within the bundle and maintaining their locations relative to their immediate neighbours over long distances. This ordering is independent of the size of the process bundle. For example, the four anterior sub-lateral cords, which are made up of only two processes, each have the same relative disposition of processes (figure 7). On the other hand, the ventral cord near the junction of the nerve ring is made up of about 170 processes; it is bilaterally symmetric in this region and the degree of order that was found can be seen by comparing the positions of bilaterally symmetrical processes on each side of the cord (figure 16b). There is a little more variability seen between the cords of different animals of the same genotype and developmental stage than between each side of the cord in a single animal. Although the order of processes in the cord is maintained over long distances, local mechanical intrusions, such as cell bodies, can disturb the ordering temporarily, but order returns away from these regions.
Processes that must have grown in opposite directions are found to be freely mixed within process bundles. The processes of PVQR and PVPR in the ventral cord, for example, must have grown up from their cell bodies in the tail, yet most of their surrounding processes, such as those of AVAL, HSNR and AVJL (figure 18b), have their cell bodies in the head and their processes must therefore have grown in opposite directions to those of PVQ and PVP. The relative positions of adjacent processes that had grown in opposite directions was fairly constant over long distances. Such an organization of processes might conceivably have arisen by rapid and sequential process growth; in other words, each process would grow along the full length of the process bundle before its neighbour growing in the opposite direction started out. A more likely explanation for these observations is that processes can insinuate themselves in between pre-existing processes in a bundle and follow along specific neighbours. The observation that the processes of PVNL in the left sub-dorsal region of the nerve ring must have grown in opposite directions, but nevertheless, ran in the same region of the nerve ring, supports this latter interpretation.
Any individual process in a bundle has a group of adjacent processes that immediately surround it at any point. We refer to such a group as the neighbourhood of the process. Neighbourhoods are generally fairly constant over the length of processes, reflecting the ordered arrangement of processes within bundles. Certain neighbours are found to be much more persistent than others, however, always remaining adjacent, whereas others move in and out of direct adjacency along the length of the process (White et al. 1983). In some instances, groups of processes are seen to be closely associated together; the most striking example of this behaviour is shown by the dendritic regions of RMD motoneurons, which are clustered around the processes of RIA (RMD-d). In this particular case there are extensive synaptic interactions between RIA and RMD, but in other cases, such as the close association of ALM and ALN on the lateral lines (ALN-d), there are no synaptic interactions between the associated processes.
FIGURE 16. Transverse sections through the anterior regions of the ventral ganglion (a) and process identification (b). The ventral cord enlarges at its anterior end and contains some of the neuropile of the cells of the ventral ganglion. Several of the processes in this region have characteristic shapes and synapses; these differences facilitate their identification. Processes from SMB flatten out and sandwich each other; AIY makes characteristic dyadic and triadic synapses onto the RIB, AIZ and RIA; FLP makes prominent synapses onto AVA; RIGL/R run near the centre of the neuropile making extensive gap junctions to each other; the processes of AIA enlarge and make several synapses to AIB and RIF. The neuropile in this region is bounded on the ventral and lateral sides by thin sheet-like processes from CEPsh. Processes from the excretory glands run along the dorsal surface of the neuropile and eventually end in the ventral regions of the nerve ring. The four cell bodies above the neuropile are always in this configuration and can be readily identified. The rest of the cell bodies in the ventral ganglion are more variable in their relative positions. *WA editors' note: PVPL and PVPR are inadvertently mislabeled in this figure. It should be PVPL on the right side of VC and PVPR on the left.
Many processes make abrupt changes of neighbourhood at certain points. The processes of AIB are closely associated with those of AIA on the ipsilateral side, but at the point where the latter terminate, on the dorsal mid-line, the processes of AIB turn and run across the process bundle. They then run for a short distance anteriorly before turning again and continuing on their trajectories round the ring; they are now in a different neighbourhood, where they run in close association with the processes of RIM (White et al. 1983). Such major changes of neighbourhood obviously have considerable functional significance for a neuron as they provide an extended set of possible synaptic partners. Perhaps, more significantly, they also facilitate direct communication between non-adjacent neighbourhoods.
In many (but not all) cases, there are external discontinuities at the transition points between neighbourhoods. The greatest numbers of neighbourhood transitions are seen to occur at the junction of two process bundles. In the region where the amphid commissure (figure 17) joins the anterior ventral cord (figure 16b), most processes from the commissure make transitions of neighbourhood (as in ASG) although some neighbours are maintained (as in AIB/AWC). The same type of behaviour occurs at the junction of the ventral cord and the nerve ring (figures 16 and 20), with some processes maintaining their neighbourhoods (see, for example, ASJ/PVQ/ASK) while others (e.g. ASH) switch. A discontinuity of a different type is seen on the dorsal mid-line or the nerve ring, which corresponds to the points where AIB, AVE and AVD make abrupt transitions of neighbourhood. In this case the discontinuity is apparently due to the termination of many processes in this region (notably the amphid receptor neurons), usually in gap junctions to their symmetrical analogues (e.g. ASJ-c). AIB and AVE are both closely associated with processes that terminate in this way in one of their neighbourhoods. In AVD, the associated processes are not obvious and it appears that the processes of AVD may have been deflected by a process emanating out of the cell body of RID (AVD-e).
Motoneurons are generally found to inhabit two neighbourhoods. One corresponds to the region where the motoneuron is predominantly or exclusively postsynaptic, usually in the interior of a process bundle, and the other is the region where NMJs are situated, at the surface of a process bundle adjacent to the basal lamina. The transitions between these neighbourhoods are not accompanied by obvious external discontinuities in most cases, except for a similar transition occurring in an adjacent motoneuron of the same class.
FIGURE 17. Diagram showing the disposition of processes within the amphid, sub-lateral and deirid commissures on each side of the animal. (See commissures in color.)
Groups of processes that are fasciculated together have been shown to share a common antigenic determinant in the leech (Hockfield & McKay 1983) and the grasshopper (Raper et al. 1983). It is possible that such antigens are neighbourhood-specific adhesion molecules. Such specific adhesion molecules, or perhaps a single ubiquitous molecule, such as CAM, which is spatially and temporally regulated (Edelman 1983), may be the basis for the close associations of groups of processes seen in the nervous system of C. elegans. It is interesting to consider the abrupt changes in neighbourhood exhibited by some neurons in the context of inter-process adhesivity. In the switches in neighbourhood that occur at process bundle junctions, it seems likely that mechanical disturbances have mixed the processes, introducing them to novel neighbours. Some of these neighbours may have high adhesive affinities for the newly introduced processes and act to guide and establish the processes in their new territory. Such a notion carries the implication that specific neighbourhoods are not uniquely attractive for a particular process, but rather that there may be several neighbourhoods in which a process could equally well reside, the one selected being dependent on the initial placement of the process in the bundle. In general there are few directed movements of neurons relative to their neighbours after they are born (Sulston 1983). It therefore seems that the initial placement of a neuron at birth is the major factor that determines which neighbourhood is finally selected out of the set of neighbourhoods in which its process could equally well reside.
The neighbourhood transitions exhibited by motoneurons seem to be mediated by factors that are intrinsic to the neuron. Other neurons, such as AVA and AVE, show a clear differentiation of their processes into regions that are both pre- and postsynaptic, and regions that are exclusively postsynaptic. In the case of motoneurons it is not clear whether there are no synapses made by the axon when it is in the interior of the process bundle because there are no suitable postsynaptic targets (muscle arms) available in the neighbourhood, or whether this region of the process is intrinsically incapable of supporting synapses. If this latter interpretation is correct, it may be that particular adhesion factors are also associated with these differentiated regions of the process. A factor that was localized in presynaptic regions that conferred an adhesive affinity with the basal lamina could, for example, serve to constrain the process to run alongside the basal lamina in these regions.
FIGURE 18. Transverse section through the ventral cord (above) and process identifications (below). The ventral cord consists of a process bundle that runs alongside a longitudinal ridge of hypodermis; the whole structure is bounded by a thin basal lamina (EL). Axons of motoneurons arrange themselves next to the basal lamina on the right-hand side of the cord in a fixed arrangement. The usual sequence of motoneuron classes from dorsal to ventral is VCn, VDn, DDn, VAn, and VBn. NMJs are made in this region (one from a VD3 is seen in this section); the motoneurons synapse through the basal lamina onto muscle arms (MA) from both left and right ventral muscle quadrants. The NMJs of a motoneuron are in a well-defined region along its process; outside this region, the process moves away from the basal lamina to the ventral regions of the process bundle. The VDn and DDn neurons are an exception in that their processes terminate abruptly outside the NMJ regions. The cell bodies of the motoneurons that innervate body muscles are arranged in a linear sequence in the ventral cord (figure 4). The ventral cord also contains the interneurons that synapse onto these motoneurons and other interneurons with little or no synaptic activity in the cord. The arrangement of processes in the cord is fairly consistent along the length of the cord, although there may be local distortions. Fingers of hypodermis (HDC) often project from hypodermal cells and run along the cord for short distances. Muscle cells have darkly staining, conical, dense bodies (DE) in the Z bands. WA editors' note: PVPL and PVPR axons are inadvertently mislabeled in this figure. It should be PVPR axon within the left side VC and PVPL axon in the right side VC (See Chapter 3 section 3.5 in R. Durbin's thesis).
FIGURE 19. Transverse section through the dorsal cord (above) and process identifications (below). The dorsal cord is similar in overall structure to the ventral cord but is much simpler, as it has fewer processes and no cell bodies. The processes in the dorsal cord are all motoneuron axones except for the processes of VDn and RMED. DAn, DEn, ASn, DDn and VDn all have processes in the dorsal cord that originate from cell bodies in the ventral cord via circumferential commissures (figure 7). RID sends a process along the length of dorsal cord from its cell body, which is situated in the dorsal ganglion.
FIGURE 20. Reconstructed cross sections of neuropile of nerve ring; (a) left lateral, (b) right lateral, (c) dorsal and (d) ventral. These drawings were obtained by reconstructing pictures of transverse sections of the nerve ring, so there are no single corresponding electron micrographs. The large outlines peripheral to the neuropile are cell bodies. The relative disposition of processes in the nerve ring is relatively constant; several processes can usually be identified directly from an electron micrograph by their morphology and position in the process bundle. The axonal regions of motoneurons are situated adjacent to the anterior inner surface of the nerve ring and synapse onto muscle arms through a basal lamina. The processes of GLR cells flatten out in this region and form a cylinder on the inside of the muscle arms. The neuropile of the ring is fairly regionalized; the amphid receptors and their associated interneurons have processes in the posterior of the ring, the neurons that make up the sub-lateral cords have their processes in the center and the mechanoreceptors tend to have processes in the anterior regions.
Circuitry
We have summarized the connectivity data or the neurons detailed in Appendix 1 into a set or connectivity diagrams (figure 21 a-r). In these diagrams, we have lumped together all members or a class and considered the connectivity or the class as a whole. Connectivity was used as one or the main criteria for grouping neurons into classes and so, by definition, all neurons within a class have the same, or very similar, patterns of connectivity to members of other classes. Thus such class groupings considerably simplify the circuit diagrams but at the expense of obscuring intraclass differences in synaptic connectivity. Such differences do not break class rules but specify which particular member of a class synapses to which particular member of another class.
The connections between classes that are shown are those that are considered to be significant. In addition, some indication has been given of the relative prominence of chemical synapses. A number of criteria were considered when making these judgements. For chemical synapses, the numbers and sizes of the synapses in a particular connection were taken into account. In marginal cases, where there were only one or a few small synapses, consideration was also given as to whether the synaptic contacts were all dyadic (with the consequent ambiguities in the identification of the functional postsynaptic partners) and whether they were present on symmetrical analogues, or corresponding cells in another animal. In the case of gap junctions, the main criteria were the area of contact and darkness of staining of the structures, and again whether they were present between analogous partners in the same or in other animals.
FIGURE 21. Circuit diagrams of nervous system. Diagrams show the pattern of connections made via gap junctions (T) and via chemical synapses (arrows) between classes of neuron. Sensory neuron classes are represented by triangles, interneurons by hexagons and motoneurons by circles. Chemical synaptic connections are graded according to their prominence on a scale of 1 to 4 (cross-hatches on arrows). Most neuron classes have been included in the diagrams; some have been included in more than one diagram for clarity.
Triangular patterns of connectivity
One of the striking features of the connectivity diagrams is the high incidence of triangular connections linking three classes. These structures may occur frequently as a consequence of the organization of the neuropile. A typical neuron in C. elegans is accessible (i.e. adjacent) to a fairly limited subset of the total complement of neurons but is fairly highly locally connected within this subset (White et al. 1983). Thus, if a neuron has synaptic contacts with two partners, these two partners must be neighbours to the neuron and therefore are likely to be neighbours themselves. It is therefore quite probable, given the high level of local connectivity, that there will be a synaptic contact between them, which will close the triangle. The abundance of triangular connections in the nervous system of C. elegans may thus simply be a consequence of the high levels of connectivity that are present within neighbourhoods.
Gap junction circuitry
Of the 104 classes of neuron in the main (i.e. non-pharyngeal) nervous system, 92 have gap junctions. Many of these classes make gap junctions to members of their own class if they are accessible to them (48 classes form such intraclass junctions). This is in marked contrast to the chemical synapses, where unambiguous synapses between members of the same class are extremely rare. Gap junctions are the presumed mediators of electrical coupling between cells, and so it seems likely that the gap junctions seen between members of a class may act to smooth discontinuities of electrical activity between adjacent class members. This may be important for classes such as the ventral cord motoneurons, for example, where marked differences in activity of adjacent motoneurons may be inimical to the smooth wave propagation required for locomotion.
Many neurons have a process that terminates at its point of contact with a process from a neuron of the same class. Most of the neurons of the amphid sensilla behave in this way, as do the DDn and VDn motoneurons of the ventral cord. In nearly all the cases where this apparent contact termination of process growth is seen, there is a gap junction at the site of contact. (The processes of RIF on the dorsal mid-line are the one striking counterexample to this general rule.) It seems possible that, in these cases, gap junctions may facilitate intercellular communication of the signals for inhibiting process extension.
Functional classification of neuron classes The simplest functional groupings of neurons that are usually made are their categorizations as either receptor neurons, interneurons or motoneurons. We have used symbols to represent these neuron types in the connectivity graphs of figure 21. Assignment of a particular class to a group is, however, not straightforward; several neuronal classes have to be assigned to more than one group, because they appear to combine two or more of these basic functions. We will go on to discuss some of the characteristics of neurons in each of these three major groupings.
Sensory receptors
The lack of electrophysiological data on any of the neurons of C. elegans makes the identification of sensory receptors and their associated modalities rather tentative. We have, however, selected a set of 39 neurons, which, on the basis of morphology and connectivity, are likely to function as sensory receptors; these have been listed in table 1.
TABLE 1. Putative sensory receptors.
The component neurons of sensilla are the neurons that are most likely to have a sensory transduction function (Ward et al. 1975). There are two general types of sensillum: those that have channels that open to the outside, exposing some or all of the neurons to the external chemical environment, and those that have no such channel. The former class is generally considered to be chemosensory and the latter, mechanosensory in function. The component neurons of sensilla are all ciliated and some of the presumed mechanoreceptors also have ciliary rootlets. There are several other classes of neurons that are not components of sensilla but which we suspect may be sensory transducers; these are also listed in table 1. The factors that have been taken as being indicative of a possible sensory function are: the presence of a cilium, the presence of a specialized, morphologically differentiated ending or the presence of a long, morphologically undifferentiated process that projects into the extremities (the tailspike or the tip of the head). In addition to these criteria, all the putative receptors should be exclusively or predominantly presynaptic.
Of the putative receptors listed in table 1, one group has a definitely known modality; another's is known with a fair degree of confidence. Laser ablation studies have shown that ALM, PLM, AVM and probably PVM transduce touch, i.e. light mechanical pressure (Chalfie & Sulston 1981). The amphid sensilla are strongly implicated as being necessary for the chemotaxis response, as several chemotaxis-defective mutants have aberrant amphidial neurones (Lewis & Hodgkin 1977).
Interneurons The interneurons in C. elegans are fairly diverse in their general organization, but some classes are conspicuous in that they are restricted in the classes of neuron with which they interact. The interneurons AIA, AIR, AIY and AIZ, for instance, receive synaptic input predominantly from the neurons of the amphid sensilla (figure 21a), whereas RIC receives its synaptic input from putative mechanoreceptors (figure 21b). Other interneurons do not show such a restriction in sensory modalities and receive synaptic input from many sources (see, for example, AVA).
The only other striking grouping that is seen in interneurones is of the classes whose synaptic outputs are directed primarily to motoneurons. These classes are AVA, AVR, AVD, AVE and PVC, which synapse onto motoneurons in the ventral cord, and RIA, which synapses onto motoneurons in the nerve ring. These interneuron classes are among the most prominent neurons in the whole nervous system. They generally have larger-diameter processes than other neurons and have many synaptic connections.
Motoneurons Each of the motoneurons in C. elegans innervates a specific group of muscle cells. This is particularly noticeable in the head region, where there is a fairly precise mapping of motoneurons onto their target muscles. Body-wall muscles are innervated by motoneurons in both the nerve ring and ventral cord. Each of these regions of neuropile contains its own unique set of motoneuron classes. The body-wall muscles can be logically divided into three regions according to the source of innervation: the head region, which receives innervation from motoneurons in the nerve ring, the neck region, which is dually innervated by motoneurons of the nerve ring and ventral cord, and the rest of the body region, which is innervated by motoneurons of the ventral cord (figure 10).
Each member of a motoneuron class in the nerve ring generally innervates muscle cells in two adjacent rows (table 2). Motoneuron classes with fourfold symmetry innervate all eight rows of muscle with no overlap, whereas motoneurons with sixfold symmetry have fields of innervation that overlap with each other by one row on each side but not across the dorso-ventral mid-line (table 2). Most of the classes of motoneuron with bilateral symmetry innervate only the lateral four rows; the exception is RIV, which only innervates ventral rows (table 2).
In addition to the intraclass circumferential mapping shown by the ring motoneurons there is also some anteroposterior mapping between classes; some motoneuron classes only innervate head muscles, some only neck muscles, while others innervate both (tables 2 and 3).
TABLE 2. Muscles innervated by motoneurons in the nerve ring. TABLE 3. Major motoneuron classes.
The muscles in the main part of the body are not so precisely mapped by motoneurons as those in the head. The ventral cord motoneurons either innervate dorsal muscles or ventral muscles (table 3), there being no finer circumferential divisions. The members of each class are evenly distributed along the length of the cord and so give rise to a longitudinal mapping onto the body muscles.
The vulval muscles are innervated by two main classes of motoneuron, VCn and HSN. The VCn motoneurons also innervate the ventral body muscles but the HSNs never do this, synapsing exclusively onto the vulval muscles. The other classes of motoneuron that innervate ventral body muscles (VAn, VEn and VDn) also have a few synapses onto the vulval muscles (figure 11); thus the HSNs appear to be the only neurons that are specific for these muscles.
The only motoneuron class that has been seen to synapse onto the set of muscles that mediate defecation is DVB, but only via a single synapse onto the intestinal muscles. The defecation muscles are all coupled together via gap junctions, so it is possible that this single synapse from DVB is the route by which defecation is controlled from the central nervous system. DVB also makes a few synapses onto body muscles.
Several motoneuron classes have long, apparently undifferentiated processes, distal to the regions where NMJs are situated, before they eventually terminate (table 3). It has been suggested in the case of the ventral cord motoneurons VA, DA, VE and DE, that these regions may function as stretch receptors (L. Eyerly and R. L. Russell, personal communication). These processes will be stretched when the body bends. This arrangement of the stretch- receptive region adjacent to the NMJ region will therefore result in body curvature's being transduced into motor activity in an adjacent region. This will mediate the translation of the region of curvature along the body. The ring motoneurons have processes that run circumferentially around the nerve ring. Two classes of motoneuron in the nerve ring have processes that leave the nerve ring distally from the region where their NMJs are situated (SME and SMD). These processes turn and run longitudinally down the sub-lateral cords. Running in these locations, these processes are ideally situated to monitor bend in the anterior body, if these processes have a stretch-transducing function. This would not be the case, however, if they ran round the nerve ring following on from their proximal regions.
Several classes of motoneuron have processes that are postsynaptic at the NMJs of other neuron classes (table 3), and have their NMJs diametrically opposite these postsynaptic regions. There is nearly always another neuron present, of the same or similar class, which has the converse arrangement of postsynaptic and presynaptic regions, i.e. it has NMJs where its partner is postsynaptic and is itself postsynaptic in the diametrically opposite region where its partner has NMJs (the DDn neurons in the Ll are notable exceptions to this generalization - White et al. 1978). This reciprocal arrangement of pairs of such neurons suggests that they may act as reciprocal inhibitors, picking up excitatory synaptic input to muscles from other classes of neuron and relaying this round to the other side of the animal as an inhibitory input to the diametrically opposite muscles, ensuring that they work in antiphase. The postsynaptic regions of these putative cross-inhibitor classes often receive a few synapses from their contralateral partners (RMD has rather more of these connections than other motoneuron classes of this type). If these synapses are inhibitory, as is assumed to be the case for the NMJs, then they could add a certain amount of positive feedback to the system. This would have the effect that when the other (i.e. non-cross-inhibitor) neuron classes are activated, the system would act as a bi-stable switch with one side activated and the other inhibited. If the cross-inhibitors have a time dependent component in their response to stimulation, then the system could oscillate, one side being activated after the other in succession.
Two classes of motoneuron that have their NMJs in the nerve ring, IL1 and URA, are also probably sensory receptors. The IL1 neurons are components of the inner labial sensilla; they may respond to mechanical stimulation at the extreme tip of the head. Presumably such a simple connection acting directly on to muscles can only mediate a simple withdrawal response. The function of URA is not clear; it is probably a sensory receptor as it is predominantly presynaptic in the ring and sends processes to the tip of the head, but the appearance and disposition of these presumed sensory endings gives no indication as to their sensory modality.
Connectivity
The availability of the complete connectivity data for a nervous system generates an almost irresistible desire to speculate extensively on the function of such a structure. We will, however, try to resist this temptation and leave such speculations for future work, when we hope that they can be backed up by corroborative experimental data. We will, therefore, try to confine our comments to the general features of the connectivity, some of which may not be obvious from the connectivity diagrams, and to the functional aspects of those parts of the circuitry for which there is some relevant experimental data.
Amphids (figure 21 a)
The neurons of the amphid sensilla have synaptic outputs that are predominantly focused onto four interneurons: AIA, AIB, AIY and AIZ. Most of the receptors that are situated in the amphid channel synapse onto the AIA-AIB pair, whereas most of the accessory neurons that are associated with the amphid sheath cells synapse onto AIY-AIZ. The amphid channel receptor, ASJ, is unusual in that it alone synapses onto none of the four main amphid associated interneuron classes, but instead synapses onto PVQ, an interneuron class that has cell bodies in the tail. PVQ also receives synaptic input from the phasmid receptor neurons, PHA, in the tail, and synapses onto AIA, thereby providing an indirect route from ASJ onto the major interneurons.
The interneurons AIA and AIB generally receive a common synaptic input from their presynaptic partners. These are usually (but not exclusively) mediated by dyadic synapses, with the closely associated processes of AIA and AIB being the postsynaptic elements. There is generally a bias to AIA, in that receptor neurons often have additional monadic synapses to AIA or dyadic synapses to AIA with an alternative co-recipient. The main synaptic output of AIA is onto AIB, and this closes the triangles made by all the neurons that synapse onto both AIA and AIB. The output from these triangular subcircuits is derived from AIB and is mainly directed to the nerve ring motoneurons, RIM, and the ventral cord interneurons, AVB. The interneurons AIY and AIZ do not make as many triangular connections as are seen on AIA and AIB, although AIY synapses onto AIZ in an analogous way to the synapse from AIA onto AIB. The main synaptic outputs of both AIY and AIZ are onto RIA interneurons, which in turn synapse onto the putative cross-inhibitor motoneurons of the nerve ring, RMD and SMD.
Several of the receptor neurons make direct synaptic contacts with some of the other major interneurons, thereby bypassing the AIA-AIB-AIY-AIZ system. Most notable of these are the connections made by ASH and ADF onto RIA and the somewhat less prominent connections made by ASH and ADL onto the ventral cord interneurons AVA, AVB and AVD.
There are several instances of receptor neurons synapsing directly onto other receptor neurons. Some of these synapses are quite striking (that of ASE onto AWC, for example) and some receptors synapse onto more than one other receptor. These receptor-receptor synapses are not peculiar to the amphid receptors as they are seen between many different classes of receptor neuron, although the amphid receptors predominantly synapse onto other amphid receptors. It seems likely that such receptor-receptor connections facilitate the modulation of the activity of one receptor by another.
Other receptors in the head and their associated interneurons (figure 21 b)
Many of the putative sensory receptors in the head, apart from those of the amphid sensilla, have connections either directly or indirectly to the five major classes of ventral cord interneuron that innervate body muscles (AVA, AVB, AVD, AVE and PVC). OLL and CEP synapse directly onto AVE; CEP and OLQ synapse onto RIC, which in turn synapses onto AVA, for example. There are also connections to the motoneurons in the nerve ring, such as the direct connections made by OLL onto SMD or the connections made to SMD and SMB by OLQ and CEP indirectly via RIC. Most of the putative sensory receptors are not exclusively postsynaptic but receive synaptic input primarily from other sensory receptors; however, these receptor-receptor connections are not as prominent as receptor-interneuron or receptor- motoneuron connections. The only receptors with a well characterized sensory modality are the touch receptors ALM, PLM and AVM (Chalfie & Sulston 1981). ALM and AVM have long, differentiated processes that run in the anterior regions of the body, whereas the processes of PLM span the posterior regions. Stimulation of the anterior neurons, by gently stroking animals with a fine hair, causes animals to move backwards; stimulation of the posterior neurons causes the animals to move forwards. Laser ablation studies have shown that these responses are primarily mediated by the connections made to AVA, AVD, PVC and AVB (Chalfie et al. 1984).
Two prominent motoneuron classes, RMD and SMD, are probable cross-inhibitors in the nerve ring. RMD receives extensive synaptic input from most of the motoneurons of the ring (including itself) at dyadic NMJs. Each of the SMD neurons has only one dendritic process that enters the NMJ region of the ring; this is postsynaptic to RME and the contralateral SMD. The dorsal and ventral RMEs (RMED and RMEV) have dendritic processes that are postsynaptic at the NMJs made by SMB. The lateral RMEs have no such processes, however, and so it is not clear whether this class should be considered to be a cross-inhibitor.
The putative cross-inhibitors of the nerve ring receive extensive synaptic input from interneurons. This is quite unlike their counterparts in the ventral cord (DDn and VDn), which are only postsynaptic to ventral cord motoneurons at NMJs. The RIP interneurons, which provide the only connection between the central nervous system and the pharyngeal nervous system, have several of the features of cross-inhibitor motoneurons; they are postsynaptic at the NMJs made by the receptors IL1 and URA, and have axonal processes that cross over to the contralateral side. It seems likely that they may act to inhibit pharyngeal pumping on receipt of an appropriate stimulus from IL1, URA or IL2.
The major source of synaptic input to the RMD and SMD cross-inhibitors comes via extensive synapses from RIA interneurons. These connections are reciprocal; the reverse connections are quite significant although not as numerous as the forward connections. RIA is one of the most prominent interneurons in the nerve ring and receives extensive synaptic input from the RIB interneurons, neurons associated with the amphid sensilla and other putative sensory receptors with no obvious modality. RIB is also a fairly prominent interneuron, which makes synaptic connections with diverse partners.
The putative receptors IL1 and URA are both fairly prominent motoneurons in the nerve ring. They behave as other motoneurons, and make quite extensive NMJs, which are also presynaptic to cross-inhibitor neurons. They also receive synaptic input from other putative receptor neurons, notably IL2 and CEP. The IL2 receptors share the same inner labial sensilla as the IL1 receptors or motoneurons, but unlike the IL1 receptors they are open to the outside and so are probably chemoreceptive.
The SAA interneurons have long, anteriorly directed, undifferentiated processes that run in the sub-lateral cords. These processes could possibly act as stretch receptors monitoring the posture of the tip of the head. The main synaptic output of SAA is directed to the major ring motoneurons, RIM, and the ventral cord interneurons, AVA. There is synaptic input from the SMB motoneurons and the VB1 ventral cord motoneurons. Thus SAA interacts with the body and the head motor systems and, given its possible head-posture transducing function, it seems likely that these interneurons could function to couple and coordinate head and body movements. Such coupling seems to occur during forward locomotion, as there are no discontinuities between head and body movements in this situation.
Motoneurons of the ventral cord (figure 21 d)
The ventral and dorsal body muscles are innervated by their own sets of motoneurons. Both sets of motoneurons have cell bodies that reside in the ventral cord (figure 4) and receive their synaptic inputs from interneurons that have processes that run along the cord. The motoneurons that innervate dorsal muscles have axons that run in the dorsal cord and join up to their cell bodies in the ventral cord via circumferential commissures (figure 7).
There are four classes of motoneuron that innervate ventral muscles (VAn, VBn, VDn, and VCn), and four that innervate dorsal muscles (DAn, DBn, DDn and ASn). Of these, the VAn and DAn classes are similar and should probably be considered to be the same class, as both have forward-directed axons and both have the same pattern of synaptic input from interneurons in the cord. In an analogous way, VBn and DBn should probably be considered as one class, as again both have the same pattern of synaptic input and the same direction of axon projection, only in this case they are posteriorly directed. All four of these classes have long, undifferentiated distal regions on their axons, in contrast to the processes of VDn and DDn motoneurons, which end abruptly at the point of contact with the process of an adjacent neuron of the same class.
The VDn and DDn motoneurons receive their synaptic input solely from the other motoneuron classes on one side of the animal, usually at dyadic NMJs, and have their own NMJs on the opposite side. On the dorsal side, DDn has NMJs and VDn is postsynaptic; on the ventral side, VDn has NMJs and DDn is postsynaptic. The VDn and DDn could again be considered as a single class; the disposition of their processes and axons suggests that they probably are cross-inhibitors. The DDn neurons have been shown to rewire in the course of larval development (White et al. 1978). In the L1 (first stage) larva their polarity is reversed from that of the adult, having NMJs on the ventral side and being postsynaptic to DAn and DEn motoneurons on the dorsal side. The DAn, DBn, SAB and DDn are the only classes of motoneuron present in the L1 ventral cord; the other classes develop post-embryonically (Sulston & Horvitz 1977).
The SAB neurons have no synaptic outputs in the adult and L4 larval stages, but in the first stage (L1) larva the three neurons of this class innervate anterior ventral body muscles (SAB-b). The only other motoneurons that are seen to innervate ventral body muscles at this stage are the putative cross-inhibitors DDn. This perhaps suggests that the SAB motoneurons may provide some excitatory inputs to the ventral body muscles during this stage. In several ways SAB neurons resemble VAn-DAn neurons. They have the same pattern of synaptic input as these classes and also have long undifferentiated distal endings to their anteriorly directed processes. These processes run in the sub-lateral cords, unlike the distal processes of VAn-DAn, which run ventrally.
The two remaining classes, ASn and VCn, are quite distinct and are less prominent with respect to their innervation of body muscles than the other classes. The ASn motoneurons innervate dorsal muscles and are somewhat similar to DAn motoneurons in morphology and synaptic input. The VCn motoneurons are primarily motoneurons for the vulval muscles (figure 11), but also innervate ventral body muscles.
There are five main classes of interneuron that provide synaptic input to the motoneurons of the ventral cord: AVA, AVE, AVD, AVE and PVC. All have cell bodies anteriorly in the lateral ganglia, except for PVC, which has its cell bodies in the lumbar ganglia in the tail. The classes AVD and AVE have identical patterns of synaptic output although they have quite different patterns of synaptic input. The processes of AVE terminate in the mid-body region, whereas the processes of all the other interneuron classes run the whole length of the ventral nerve cord. AVA, AVD and AVE make chemical synapses onto the VAn-DAn motoneurons; AVA also makes gap junctions to them. The dorsal motoneuron class, ASn, has all the classes of synaptic partner that VAn-DAn motoneurons have, and indeed makes gap junctions with them, but it receives an additional chemical synapse from AVE.
The VBn-DBn motoneurons are predominantly innervated by gap junctions from AVE and chemical synapses from PVC together with a few chemical synapses from DVA. Laser ablation experiments have demonstrated that, in the first stage larva, the DEn motoneurons are necessary for forward locomotion (backward-propagating body waves), and the DAn motoneurons are necessary for backward locomotion (forward-propagating body wave) (Chalfie et al. 1984). Because of their similar structure and identical patterns of synaptic input, it seems likely that VAn motoneurons have similar functions to DAn motoneurons, and likewise VBn motoneurons have similar functions to DBn motoneurons. Considering the sources of synaptic input to these classes of motoneuron, it seems likely that the AVB-PVC interneurons are used for forward movement and the AVA-AVD-AVE interneurons are used for backward movement. There is some evidence for this from laser ablation studies (Chalfie et al. 1984).
Circuitry associated with neurons in the tail (figure 21 e)
The tail region of C. elegans contains a number of classes of receptor neuron, interneuron and motoneuron that are specific to this region. Most of these neurons project into the neuropile of the pre-anal ganglion, which is situated at the posterior extremity of the ventral cord. In general, synapses made by neurons in the tail are smaller and less numerous than those seen in the nerve ring or anterior ventral cord. Some classes of neuron, such as PVT, PVW and PDB, make very few synaptic contacts. The major interneurons in the tail circuitry are the ventral cord neurons, AVA, AVD and PVC, and two interneuron classes with cell bodies in the tail, DVA and LUA.
The tail has two pairs of sensilla, the phasmids and the posterior deirids. The phasmids are probably chemosensory, as their component neurons are open to the outside in a similar arrangement to the neurons of the amphid sensilla. There are two neurons in each sensillum, PHA and PHB. PHA is unusual; virtually all its synaptic output is directed onto the other phasmid neuron, PHB. This in turn synapses mainly onto AVA and PVC.
The posterior deirid sensilla are similar in structure to the anterior deirids, and both have been shown to contain the neurotransmitter dopamine (Sulston et al. 1975). The cell bodies of the single receptor neuron (PDE) and the accessory cells of the sensilla are situated in the lateral, mid-posterior regions of the body. The synaptic output of PDE is quite different from that of the anterior deirid receptor neuron, ADE; its main postsynaptic partner is DVA. The putative receptor neuron PVM has a cell body in the right-hand posterior lateral ganglion and has a differentiated ultrastructure that is very similar to that of the anterior touch receptor neuron, AVM (Chalfie & Sulston 1981). Its synaptic output is quite different from that of AVM, however, being directed mainly to PDE. This neuron does not seem to be involved in the touch response (Chalfie et al. 1984) as is AVM.
The posterior body of the hermaphrodite tapers down into along thin tailspike. Seven classes of neuron have long, undifferentiated processes that run nearly to the end of this tails pike (AVG, ALN, PLN, PHC, PVR, PLM and PDB). It seems likely that these neurons are sensory and that the tailspike is, in fact, a large sense organ, although it does not have the sheath and socket cells that are components of sensilla. The neurons of the tailspike are quite diverse in their synaptic connections. PHC has short processes and synapses predominantly onto DVA and PVC; PVR has a process that traverses the length of the ventral cord and synapses onto AVB and RIP in the nerve ring. AVG is the only tailspike class that does not have a posteriorly located cell body; it has a single, rather large cell body in the retrovesicular ganglion. The main synaptic output of AVG seems to be via extensive gap junctions to the two RIF interneurons also situated in the retrovesicular ganglion.
The other classes of neuron with processes in the tailspike, ALN, PLN and PLM, probably have a sensory function in other regions as well as in the tails pike. PLM are the posterior touch neurons and span the whole of the posterior region of the body. ALN and PLN are two classes that have processes that run alongside, and are closely associated with, the transducing regions of the processes of ALM and PLM respectively. They project into the nerve ring and it seems probable that they are also involved with the touch system in some way. The motoneuron PDB has a proximal process that runs into and oUt of the tailspike en route from the ventral to the dorsal cord. No synaptic input is seen onto this neuron; however, it makes a few NMJs onto dorsal body muscles. It is possible that, in contrast to other motoneurons with long distal processes, the long proximal process of PDB may have some transducing function in the tailspike. PDA is another single motoneuron like PDB; both have cell bodies situated in the pre-anal ganglion. PDA also innervates dorsal muscles but sends its process to the dorsal cord by a more direct route via a lumbar commissure. It receives some synaptic input from the interneuron/motoneuron DVB. The only synaptic input to the defecation muscles is provided by DVB, which therefore (presumably) controls defecation. PDA may mediate the contractions of the posterior body, which are associated with defecation (Crofton 1966), via its connection with DVB.
The ventral cord motoneurons, DA8, DA9 and VA12, have rather different patterns of synaptic connections from the more anterior members of their classes. Although they still retain the synaptic inputs from AVA and AVD that are characteristic of these classes, DA9, DA8 and VA12 have several additional sources of synaptic input: VA12 from PHC; DA9 from PHC and PHB; and DA8 from DVB. In addition VA12 synapses onto DB7, DA8 and DA9. None of the other VAn motoneurons is seen to synapse convincingly onto other motoneuron classes except VDn and DDn, and So this feature is probably indicative of an intrinsic difference between VA12 and the other VAns. The synaptic inputs from PHC and PHB, on the other hand, may be restricted to the posterior members of the VAn and DAn classes simply because of the limited extent of the axons of PHB and PHC in the ventral cord. This would not necessarily require an intrinsic difference in DA8, DA9 and VA12 compared with the other members of their class.
The egg-laying circuitry (figure 21 f)
The vulval and uterine muscles are predominantly innervated by two classes of motoneuron, HSN and VCn. The VCn motoneuron class has six members, which are distributed along the central regions of the ventral cord. They synapse onto ventral body muscles as well as vulval muscles. The only significant synaptic input that was seen on to them comes from HSN. These synapses are in close proximity to the NMJs made by the VCns onto the vulval muscles, suggesting that they could perhaps be mediating a presynaptic inhibition of the VCns. Various pharmacological agents, including acetylcholine agonists, serotonin analogues and an octopamine blocking agent, have been shown to stimulate egg laying (Horvitz et al. 1984).
Laser ablation experiments have shown that the HSN neurons are essential for egg laying (Trent et al. 1983). The circuitry associated with HSN is rather ambiguous. It is predominantly presynaptic and only receives a few synapses back from its postsynaptic partner, BDU, and a single synapse from each PLM. This type of behaviour suggests that HSN is not simply a motoneuron but may have some sensory transduction function that provides the primary signal for the activation of the vulval muscles. There is, however, no obvious feature of its structure which suggests such a function.
The same arguments can be applied to the VCn neurons because of their apparent lack of presynaptic partners, although it is not yet known whether the VCns are essential for egg laying. Another possibility is that the main inputs for HSN and VCn come via humoral neurotransmitters rather than by focal synaptic contacts. The sensory integration required to determine the appropriate moment for egg laying could then be executed in other regions of the nervous system, with no morphologically distinguishable connections being made to the vulval muscle motoneurons.
There are, perhaps, two fundamental questions in the field of neurobiology: how neurons organize themselves during development into specifically interconnected networks, and how such a network functions. A knowledge of the detailed structure of a nematode's nervous system does not in itself provide any answers to these questions, but it does at least provide a framework within which it is possible to pose rather more specific questions.
The development of a nervous system can be divided into three separate phases. The first is the generation of a group of differentiated neurons; the second is the outgrowth and guidance of processes from these neurons and the third is the establishment of connections between processes. The structural data on the nervous system provides information that is most pertinent to the last two phases. This is because the final structure represents the ultimate consequences of the execution of these two processes. We will go on to discuss how these two developmental processes, together with the question of nervous system function, may be further explored in C. elegans.
Process placement One of the most striking features of the nervous system of C. elegans is the precision with which processes are positioned relative to their neighbours within process bundles. Synaptic contacts are made en passant between adjacent processes; the set of possible synaptic partners that a neurone may have is therefore limited to the set of processes that are neighbours. Given the unbranched nature of nematode neurons, this set is usually a relatively small subset of the total complement of neurons that make up the nervous system. Within this neighbourhood, however, neurons are fairly highly connected, making connections to nearly half their neighbours on average (White et al. 1983). Furthermore, there is circumstantial evidence that this level of connectivity may be independent of neighbourhood, i.e. that a given neuron may make synaptic connections to more or less the same percentage of its neighbours no matter what class they may be (White et al. 1983). Thus process placement must be a major determinant in the establishment of the patterns of connectivity within the nervous system of C. elegans.
It seems likely that there may be two aspects of process placement: substrate guidance of pioneering processes to establish process tracts (Berlot & Goodman 1984), and the positioning of processes relative to their neighbours within bundles once process tracts have become established. A distinctive feature of the organization of processes within bundles is the close associations that are seen between specific processes, or between a process and the basal lamina. Such associations are probably the consequence of selective adhesive affinities between the associating entities. Given the probable importance of selective adhesivity in determining connectivity, it is worth considering, within the context of the nematode's nervous system, how such phenomena may be further investigated.
Many behavioural mutants have been isolated in C. elegans; it is likely that most of their phenotypes are the consequence of alterations in the nervous system. It is also likely that some of these alterations could take the form of misplaced processes. Up to now, relatively few behavioural mutants have been analysed at the ultrastructural level. This is mainly because of the considerable effort that is required to reconstruct a significant portion of the nervous system from electron micrographs. Recently, staining techniques have been developed that allow the visualization of specific processes or process bundles in whole mounts of C. elegans when viewed with the light microscope. In one of these techniques, sensory process tracts are labelled by dye filling (Hedgecock et al. 1984). In another, processes of certain neuron classes are labelled with monoclonal antibodies and viewed by immunofluorescence in whole mounts (Okamoto & Thomson 1984). Such techniques will facilitate the pre-screening of behavioural mutants for those that have abnormalities in process placement. Selected mutants may then be subjected to a full ultrastructural analysis.
With the dye uptake technique, certain mutants have been found to have abnormal projections from sensory receptors (Hedgecock et al. 1984); such mutants could be candidates for substrate guidance. The defects in these mutants could either be located in the neurons, or in the substrate upon which they grow. It may be possible to distinguish between these two possibilities by means of mosaic analysis (Herman 1984).
Of the mutants that have been analysed by serial section reconstruction, one (unc-30) has been found to have misplaced processes on the VDn and DDn motoneuron classes (J. G. White, S. Brenner & R. Durbin, unpublished observations). The disposition of the processes of the other motoneuron classes in the ventral cord appears normal. It seems possible that such a mutant could be defective in the class-specific expression of an adhesion factor. The molecular analysis of genes that affect process placement may provide a route to an eventual understanding of the function and deployment of region-specific adhesion molecules. Another route to the same end may be taken by directly looking for putative adhesion molecules. Candidate molecules would be expected to be common to a group of processes that are closely associated together. Such a molecule could be sought either directly by using antibodies, or indirectly by looking for species of messenger RNA that show the appropriate neuronal distribution.
Synaptic specificity Although we have played down the role of synaptic specificity in the generation of the pattern of connections within the nervous system of C. elegans to a certain extent, it is clear that there has to be some level of specificity. On average, a neuron is presynaptic to about 15% of its neighbours (unpublished observations). The subset of neighbours that are postsynaptic to a given neuron is fairly constant from animal to animal, and So is presumably actively selected. It is likely that synaptogenesis is initiated by a cell-cell recognition event. Such an event may involve the binding of a surface receptor molecule on one cell to a matching 'label' molecule on another cell. If all cell classes had single distinguishing label and receptor types, then the set of synaptic partners of a given cell class could never intersect with that of another. Such intersections are, in fact, the general rule in the nervous system. Therefore, if such a label-receptor system is the basis of synaptic specificity, then the labels (and/or receptors) have to be arranged combinatorially.
It is probably not reasonable to assume that the pattern of connections seen between processes in a particular neighbourhood is solely the consequence of the intrinsic specificities of the neurons involved. There are suggestions that interactions between synapses may act to modify certain patterns of synaptic connection that might otherwise form as a consequence of specific neuron-neuron recognition. There are slight differences in connectivity between the dorsal and ventral members of the classes SMB, SAA, OLQ and RMD. These differences are manifested as reciprocal substitutions of gap junctions for chemical synapses and chemical synapses for gap junctions. This behaviour may suggest that there are interactions between these types of connection in these circumstances, and that these interactions result in a mutual exclusivity of chemical synapses and gap junctions.
We have used the criteria of morphology and connectivity to define the 118 classes of neuron that have been described. Given that a particular neuron can only select synaptic partners from its neighbourhood, it is probable that there are classes that we have defined that have the same intrinsic synaptic potential; in other words, if placed in the same neighbourhood they would select the same subset of neighbours as synaptic partners. Therefore, the number of classes that we have defined (118) is almost certainly an overestimate of the number of neuron types that are intrinsically different in their specificities. It is strongly suspected, on the basis of morphology, that AQR and PQR are members of a single class, as are ALM and PLM, ALN and PLN, and AVM and PVM. It is probable that there are other class equivalences that are not so obvious, particularly among the interneurons, which often do not have distinguishably different morphologies. It may be possible to identify such 'superclasses' by a neighbourhood analysis. If the neighbourhoods from two classes are compared and common neighbours are identified, then it is possible that the two classes may be members of a superclass, if the pattern of synaptic connections made to the common neighbours is the same in each case. By considering all pairwise combinations of classes, and then reiterating the process considering all members of putative superclasses as equivalent, it may be possible to arrive at a logically consistent set of superclasses. These superclasses will define groups of cells that have intrinsically identical synaptic specificities. Such an endeavour may not just be an idle intellectual exercise, as a knowledge of such 'supergroups' could facilitate the identification of mutants that have altered labels or receptors. Such mutations would be expected to have pleiotropic consequences, affecting all the members of a supergroup. Thus mutants that affect connectivity of all the members of a particular supergroup are candidates for mutants with altered labels and/or receptors. An analysis of such mutants may provide a possible route towards an understanding of the molecular basis of synaptic specificity.
Nervous system function The relative simplicity of the structure of the nervous system of C. elegans provides a challenge to determine how it functions. The main disadvantage of this nervous system from the point of view of functional studies is that the small size of the component neurons precludes the use of electrophysiological recording techniques. Such techniques can, however, be used with Ascaris. There are considerable homologies between the ventral cord motoneurons of Ascaris and C. elegans (Stretton et al. 1978); more recently, similar homologies have been seen in the interneurons of the retro-vesicular ganglion (Donmoyer, Angstadt and Stretton, personal communication). The neurotransmitter dopamine has been shown to be present in the same classes of cells in the two animals (Sulston et al. 1975). It seems likely that such structural and biochemical similarities may indicate an underlying functional similarity, justifying the extrapolation of data obtained from one animal to the other. Electrophysiological studies on homologous cells in Ascaris suggest that the DAn, DBn, and ASn motoneurons of C. elegans are excitatory, whereas the DDn and VDn motoneurons are inhibitory (Johnson & Stretton 1980). Further work may yield information about the role of the interneurons of the ventral cord in activating the motoneurons.
The functional aspects of the nervous system of C. elegans may be studied directly by characterizing the behavioural consequences of specific lesions in the nervous system. Lesions may be produced by laser microsurgery (Sulston & White 1980), a technique that is capable of removing any cell or small group of cells within the nervous system. As an alternative, use may be made of lesions produced as a consequence of mutations. For example, one mutant, unc-30, specifically affects the organization of the VDn and DDn motoneurons in the ventral cord, leaving the other motoneuron classes relatively unaffected (J. G. White, S. Brenner & R. Durbin, unpublished observations). This mutant is uncoordinated in forward and backward locomotion. When stimulated by a tap on the head, instead of backing away, these animals shorten by simultaneously activating both their ventral and their dorsal muscles. This behaviour is what one would predict if cross-inhibition between the dorsal and ventral sides were lacking. This observation reinforces the suggestion, originally made on morphological criteria, that the VDn and DDn classes function as cross-inhibitors.
The combined techniques of laser microsurgery, mutants and tests for drug responsiveness have been used to produce detailed models for the function of the circuitry associated with the touch response (Chalfie et al.1984), and the circuitry that controls egg-laying (Horvitz et al. 1984). Other areas of the nervous system should be equally amenable to such methodologies, particularly the chemosensory system. This system is particularly attractive, as the chemotactic response has been characterized (Ward 1973; Dusenbery 1974) and many mutants that are defective in chemotaxis have been isolated (Dusenbery et al. 1975; Lewis & Hodgkin 1977).
Neurons
Appendices
APPENDIX 1. CONNECTIVITY DATA In this section, all the detailed connectivity data for each of the neuron classes are presented. The neuron classes are arranged in alphabetical order; the data for each class are fairly self-contained. Some classes have been grouped together because they share many common features; PLM is listed with ALM, PLN with ALN, PVM with A V M and PQR with AQR. The data that are presented were derived primarily from three reconstructed animals; the N2T series, the N2U series and the JSE series. Together these series covered the whole of the animal except for a region in the posterior body (figure A 1). This region was covered by a partial reconstruction of a male (N2Y series). Data from this animal provided information on the neurons of the posterior lateral ganglia and the motoneurons of the posterior ventral cord. The neuropile of the nerve ring and anterior ventral cord was also reconstructed from an L4 larva (JSH series, figure A 1). These data were mainly used as a check on the N2U reconstructions, which covered this region and are not shown, except in the case of RMF, where there was a significant difference between the two series.
Neuron topographies are shown in semidiagrammatic form for simplicity in presentation. Processes of neurons in C. elegans have few, if any, branches and tend to run in parallel process bundles. It is therefore possible to give a reasonably accurate impression of their three- dimensional structure by means of such diagrams. Neurons that inhabit the regions of the nerve ring and anterior ventral cord are plotted out in diagrams on templates of the form shown in figure A 2. Similarly, neurons that have processes in the posterior ventral cord are plotted out in diagrams on templates of the form shown in figure A 3. Additional diagrams show the disposition of the cell bodies and processes of the class members within the animal, as seen from a lateral viewpoint. The nerve ring or anterior ventral cord diagrams are drawn as if from a dorsal viewpoint of an animal in which the nerve ring has been flattened so as to lie in the same plane as the ventral cord. The diagrams of neurons in posterior regions are again drawn from a dorsal viewpoint, but in this case an imaginary cut has been made along the dorsal mid-line and the animal opened out flat so that the ventral mid-line runs along the centre of the diagrams (figure A 3).
Processes that run in the regions covered by these types of diagram have been drawn out with all their synaptic connections listed. Synaptic connections mediated by chemical synapses are depicted by arrows. The direction in which the arrow points relative to the process indicates whether the process is presynaptic or postsynaptic for that particular contact. Synaptic contacts in which the process is one of several that are postsynaptic to a single presynaptic element are marked with an asterisk. All possible postsynaptic partners of contacts in which the process is presynaptic are shown. Gap junctions only appear between two elements and are marked with a T; no directionality is implied.
Certain synaptic connections have additional labels. These labels refer to a set of electron micrographs, which illustrate these connections. Many illustrations were taken from the JSH series because of the better quality of the pictures that were obtained from this series. Although the diagrams refer to connections seen in the other series, it was nevertheless possible to use these illustrations, because in most cases synaptic connections equivalent to those indicated in the diagrams could be found in the JSH series. References to illustrations of synaptic contacts are made by an index letter. These refer to the set of illustrations that is associated with the neuron class currently under discussion. If the index letter is preceded by an asterisk then the index letter refers to the set of illustrations associated with the class being referenced.
The two diagram formats described above do not cover the central body region, particularly the region of the ventral cord in which there are many synaptic contacts. Data from this region are presented in two ways: either as a table of synaptic contacts, in the case of interneurons which have processes that enter the region, or as individual diagrams, for motoneurons that are totally contained within the region. The motoneurons of the ventral cord have up to thirteen members in each of the classes, compared with a maximum of four members for all the other neuron classes in the animal. Only one 'typical' member of each of the ventral cord motoneuron classes is plotted, together with any atypical members that there may be in the class.
FIGURE A 1. The regions covered by the five separate reconstructions. The N2T, N2U and JSE series were adult hermaphrodites, the JSH series was an L4 larva and the N2Y series was an adult male.
FIGURE A 2. Diagram of the projection and template used for the plots of processes that run in the nerve ring and ventral cord. The nerve ring has been flattened out to lie in the same plane as the ventral cord, so that the posterior face of the nerve ring and the dorsal face of the ventral cord are directed out of the page. The shaded region indicates the extent of the neuropile in these regions. The isthmus of the pharynx passes through the hole in the middle of the nerve ring. The disposition of the major process tracts that join this region of neuropile are shown.
FIGURE A 3. Diagram of the projection and template that is used for the plots of processes in the tail region. This is a dorsal view of the projection obtained by making an imaginary cut along the dorsal mid-line and then opening and flattening the animal. The outlines indicate the dispositions of the process tracts and ganglia. The rectum passes through the hole in the middle.
APPENDIX 2. REMAINING AMBIGUITIES The reconstructions that we have described were done piecemeal, using data obtained from several animals. This was because of the difficulty of serial sectioning a complete individual. Inevitably there were some consequential problems in the identification of equivalent processes in different animals. These problems could, for the most part, be resolved, because of the reproducibility of relative process positions within bundles and the consistent synaptic behaviour of a given process. There are, however, a few remaining ambiguities, notably concerning process identification in the posterior ventral cord. This region was covered by the N2Y series, which was derived from an adult male. The male has significantly more processes running in its ventral cord than the hermaphrodite. These extra processes arise from male-specific neurons in the tail ganglia (Sulston et al. 1980). Their presence made it difficult to identify some of the processes from neurons common to both sexes, particularly those that were rather featureless, with few characteristic synapses. We have listed the cases for which these problems exist.
DVB and DVC
The processes from these two neurons are always closely associated. They sometimes twist round each other and so cannot be distinguished by their relative positions. We cannot, therefore, be sure whether the cell bodies labelled DVB and DVC connect to the processes labelled DVB and DVC in the nerve ring or whether they have been crossed over. We have chosen the interpretation that is shown on the basis of synaptic criteria, but these are not particularly compelling.
ALA, CAN and PVD
These neuron classes have processes that run together alongside the two lateral arms of the excretory canal. They have not been followed along the length of the animal, although they have been sampled at intervals along their length. The three processes make virtually no synaptic connections along the canal and look rather similar. Two of the processes end at about the level of the anus; the third enters the lumbar ganglion on each side, where it makes a few synapses onto PVC. This process has been tentatively assigned to ALA, but it could equally well belong to either of the other classes.
PVW, PQR and PVT
These classes have cell bodies in the lumbar ganglia and the pre-anal ganglion; they send out processes, which project anteriorly up the ventral cord. Processes from these neurons have not been positively identified in the N2Y series and are not present in the anterior ventral cord (i.e. they could not be accounted for in the N2U series). It therefore seems likely that they terminate somewhere in the posterior ventral cord. The process of PQR appeared to be petering out at the end of the JSE series and was almost certainly about to end. The same cannot be said of PVT, however. The single neuron of this class had a large cell body in the pre-anal ganglion with a single, substantial, anteriorly directed process. This process had few synaptic contacts and was still going at the anterior extent of the JSE series. It seems rather surprising that such a process should end, presumably without making any significant synaptic contacts. An alternative interpretation is that this process divides somewhere in the posterior cord and becomes the two processes currently assigned to PVNL/R in the anterior cord. The location of the process of PVT in the cord is consistent with this interpretation. It would mean, however, that in this case PVNL/R would have to terminate in the posterior cord.
RID, PDA and PDB
These three classes have processes in the dorsal cord, which have not been completely followed. RID sends a process into the anterior end of the cord; a process that looks similar has been identified in the posterior dorsal cord. It therefore seems likely that the dorsal process of RID spans the length of the cord. The process of PDA and PDB enter the dorsal cord near its posterior extremity. They project anteriorly but have not been identified in the anterior cord.
APPENDIX 3. Name Equivalences In some previous publications on C. elegans neuroanatomy, different systems of nomenclature have been used. We have included a list of equivalences to facilitate cross-referencing between these papers.
We would like to thank our colleagues who, over the years, have offered advice and encouragement for this work. We would particularly like to mention Donna Albertson, Martin Chalfie, Richard Durbin, Edward Hedgecock, Robert Horvitz and John Sulston for the many stimulating discussions that we have had together, and also Donna Albertson, Leon Nawrocki and John Sulston for reading and commenting on the manuscript.
References
Albert, P. S. & Riddle, D. L. 1983 Developmental alterations in the sensory neuroanatomy of the C. elegans dauer larva. J. comp. Neurol.219, 416-481.
Albertson, D. G. & Thomson, J. N. 1976 The pharynx of Caenorhabditis elegans. Phil. Trans. lL Soc. Lond. B 275, 299-325.
Bennett, M. L. V. 1977 Electrical transmission: a functional analysis and comparison to chemical transmission. In The handbook of physiology. Section 1: the nervous system. (ed. J. M. Brookhart & V. B. Mountcastle), pp. 357-416. Bethesda, Maryland: American Physiological Society.
Berlot, J. & Goodman, C. 1984 Guidance of peripheral pioneer neurones in the grasshopper: an adhesive hierarchy of epithelial and neuronal surfaces. Science, Wash.223, 293-295.
Bird, A. F. 1971 The structure of nematodes. New York: Academic Press.
Boycott, B. B. & Dowling,J. E. 1969 Organization of the primate retina: Light microscopy. Phil. Trans. R . Soc. Lond. B 255, 109-184.
Braitenberg, V. & Atwood, R. P. 1958 Morphological observations on the cerebellar cortex. J. comp. Neurol. 109, 1-34.
Brenner, S. 1974 The genetics of Caenorhabditis elegans. Genetics, Princeton 77, 71-94.
Cajal, R. 1972 Histologie du systeme de l'homme et des vertibris. Madrid: C.S.I.C.
Cassada, R. C. & Russell, R. L. 1975 The dauer larva, a postembryonic developmental variant of the nematode Caenorhabditis elegans.Devl Biol.46, 326-342.
Chalfie, M. & Sulston, J. 1981 Developmental genetics of the mechanosensory neuron of Caenorhabditis elegans.Devl Biol.82, 358-370.
Chalfie, M., Sulston, J. E., White, J. G., Southgate, E., Thomson, J. N. & Brenner, S. 1984 The neural circuits for touch sensitivity in Caenorhabditis elegans. J. Neurosci. 5, 956-964.
Chalfie, M. & Thomson, J. N. 1982 Structural and functional diversity of neuronal microtubules of Caenorhabditis elegans. J. Cell Biol.93, 15-23.
Chitwood, B. G. & Chitwood, M. B. 1974 Introduction to nematology. Baltimore: University Park Press.
Crofton, H. D. 1966 Nematodes. London: Hutchinson.
Culotti, J. G. & Russell, R. L. 1978 Osmotic avoidance defective mutants of the nematode, Caenorhabditis elegans. Genetics, Princeton 90, 243-256.
Dowling, J. E. & Boycott, B. B. 1966 Organisation of the primate retina: electron microscopy. Proc. R. Soc. Lond. B 166, 80-11.
Dusenbery, D. B. 1974 Analysis of chemotaxis in the nematode Caenorhabditis elegans by countercurrent separation. J. exp. Zool.188, 41-47.
Dusenbery, D. B., Sheridan, R. E. & Russell, R. L. 1975 Chemotaxis defective mutants of the nematode, Caenorhabditis elegans. Genetics, Princeton 80, 297-309.
Eccles, J. C., Ito, M. & Szentigothai, J. 1967 The cerebellum as a neuronal machine. Berlin: Springer-Verlag.
Edelman, G. M. 1983 Cell adhesion molecules. Science, Wash. 219, 450-457.
Goldschmidt, R. 1908 Das Nervensystem von Ascaris lumbricoides und megalocephela, I. Z. wiss. Zool. 90, 73-136.
Goldschmidt, R. 1909 Das Nervensystem von Ascaris lumbricoides und megalocephela, II. Z. wiss. Zool. 92, 306-357.
Hall, D. H. 1977 The posterior nervous system of Caenorhabditis elegans. Ph.D. thesis, California Institute of Technology.
Hedgecock, E. M. & Russell, R. L. 1975 Normal and mutant thermotaxis in the nematode, Caenorhabditis elegans. Proc. natn. Acad. Sci. U.S.A.72, 4061-4065.
Hedgecock, E. M., Culotti, J. G., Thomson, J. N. & Perkins, L.A. 1985 Axonal guidance mutants of Caenorhabditis elegans identified by filling identified neurons with fluorescein dyes. Devl Biol. 111, 158-170.
Herman, R. K. 1984 Analysis of genetic mosaics in Caenorhabditis elegans. Genetics, Princeton 108, 165-180.
Hirsh, P., Oppenheim, D. & Klass, M. 1976 Development of the reproductive system of Caenorhabditis elegans. Devl Biol.49, 200-219.
Rockfield, S. & McKay, R. 1983 Monoclonal antibodies demonstrate the organisation of axons in the leech. J. Neurosci.3, 369-375.
Hodgkin, J. A. 1983 Male phenotypes and mating efficiency in Caenorhabditis elegans. Genetics, Princeton 103, 43-64.
Horvitz, H. R., Trent, C., White, J. G. & Sulston, J. E. 1986 Structure and function of the egg-laying system of the nematode Caenorhabditis elegans. (In preparation.)
Jan, Y. N., Bowers, C. W., Branton, D., Evans, L. & Jan, L. Y. 1983 Peptides in neural function: Studies using frog autonomic ganglia. CSH Symp. quant. Biol.48, 363-374.
Johnson, C. D. & Stretton, A. 0. W. 1980 Neural control of locomotion in Ascaris: Anatomy, electrophysiology and biochemistry. In Nematodes as biological models (ed. B. Zukerman) New York and London: Academic Press.
Kimble, J. 1981 Alterations in cell lineage following laser ablation of cells in the somatic gonad of Caenorhabditis elegans. Devl Biol.87, 286-300.
Lewis, J. A. & Hodgkin, J. A. 1977 Specific neuroanatomical changes in chemosensory mutants of the nematode, Caenorhabditis elegans. J. comp. Neural.172, 489-509.
Nelson, F. K., Albert, P. S. & Riddle, D. L. 1983 Fine structure of the Caenorhabditis elegans secretory-excretory system. J. Ultrastruct. Res. 82, 156-171.
Okamoto, H. & Thomson, J. N. 1984 Monoclonal antibodies which distinguish certain classes of neuronal and supporting cells in the nervous system of C. elegans. J. Neurosci.5, 643-653.
Pappas, G. D. & Waxman, S. G. 1977 Structure and function of synapses. New York: Raven Press.
Raper, J. A., Bastiane, M. & Goodman, C. S. 1983 Pathfinding by neural growth cones in grasshopper embryos. J. Neurosci. 3, 31-41.
Reynolds, E. S. 1963 The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol.17, 208-213.
Riddle, D. L., Swanson, M. M. & Albert, P. S.1981 Interacting genes in nematode dauer larva formation. Nature, Lond. 290, 668-671.
Rosenbluth, J. 1965 Ultrastructural organisation of obliquely striated muscle fibres in Ascaris lumbricoides. J. Cell Biol.25, 495-515.
Stevens, B. J. & White, J. G. 1979 Computer reconstruction of mitochondria from yeast. Meth. Enzym.56, 718-728.
Stretton, A. O. W., Fishpool, R. M., Southgate, E., Donmoyer, J. E., Walrond, J. P., Moses, J. E. & Kass, I. S. 1978 Structure and physiological activity of the motoneurones of the nematode Ascaris. Proc. natn. Acad. Sci.U.S.A.75, 3493-3497.
Sulston, J. E. 1983 Neuronal cell lineages in the nematode Caenorhabditis elegans. CSH Symp. quant. Biol.48, 443-452.
Sulston,J . E., Albertson, D. G. & Thomson, J. N. 1 1980 The Caenorhabditis elegans male: Postembryonic development of non gonadal structures. Devl Biol.78, 542-576.
Sulston, J., Dew, M. & Brenner, S. 1975 Dopaminergic neurons in the nematode, Caenorhabditis elegans. J. comp. Neural. 163, 215-226.
Sulston, J. E. & Horvitz, H. R. 1977 Post-embryonic lineages of the nematode, Caenorhabditis elegans. Devl Bioi.56, 110-156.
Sulston, J. E., Schierenberg, E., White, J. G. & Thomson, J. N. 1983 The embryonic cell lineage of the nematode, Caenorhabditis elegans. Devl Bioi.100, 64-119.
Sulston, J. E. & White, J. G. 1980 Regulation and cell autonomy during postembryonic development of Caenorhabditis elegans. Devl Bioi. 78, 577-597.
Swanson, M. M., Edgley, M. L. & Riddle, D. L. 1984 In Genetic maps 1984 (ed. S.J. O'Brien), pp. 286-300. New York: Cold Spring Harbor Laboratory.
Trent, C., Tsung, N. & Horvitz, H. R. 1983 Egg-laying defective mutants of the nematode, Caenorhabditis elegans. Genetics, Princeton 104, 619-647.
Wassle, H., Peichl, L. & Boycott, B. B. 1981 Dendritic territories of cat retinal ganglion cells. Nature, Lond. 292, 344-345.
Ward, S. 1973 Chemotaxis by the nematode, Caenorhabditis elegans: ldentjfication of attractants and analysis of the response by the use of mutants. Proc. natn. Acad. Sci. U.S.A. 70, 817-821.
Ward, S., Thomson, N., White, J. G. & Brenner, S. 1975 Electron microscopal reconstruction of the anterior sensory anatomy of the nematode, Caenorhabditis elegans. J. comp. Neurol.160, 313-338.
Ware, R. W., Clark, C., Crossland, K. & Russell, R. L. 1975 The nerve ring of the nematode, Caenorhabditis elegans. J. comp. Neurol.162, 71-110.
Waterston, R. H., Thomson, J. N. & Brenner, S. 1980 Mutants with altered muscle structure in Caenorhabditis elegans. Devl Bioi.77, 271-302.
Weiss, P. 1934 In vitro experiments on the factors determining the course of the outgrowing nerve fibre. J. exp. Zool.68, 393-448.
White, J. G. 1974 Computer aided reconstruction of the nervous system of C. elegans. Ph.D. thesis, University of Cambridge.
White, J. G., Albertson, D. G. & Aness, M. A. R. 1978 Connectivity changes in a class of motoneurone during the development of a nematode. Nature, Lond. 271, 764-766.
White, J. G., Southgate, E., Thomson, J. N. & Brenner, S. 1976 The structure of the ventral cord of Caenorhabditis elegans. Phil. Trans. R. Soc. Lond. B 275, 327-348.
White, J. G., Southgate, E., Thomson, J. N. & Brenner, S. 1983 Factors that determine connectivity in the nervous system of Caenorhabditis elegans. CSH Symp. quant. Bioi.48, 633-640.
Wright, K. A. 1980 Nematode sense organs. In Nematodes as biological models (ed. B. Zukerman), pp. 237-293. New York: Academic Press.
Table Guide - by WormAtlas Editors
Click picture for higher resolution image
Web adaptation, Thomas Boulin, for WormAtlas, 2002. Updated by Laura A. Herndon, 2014.

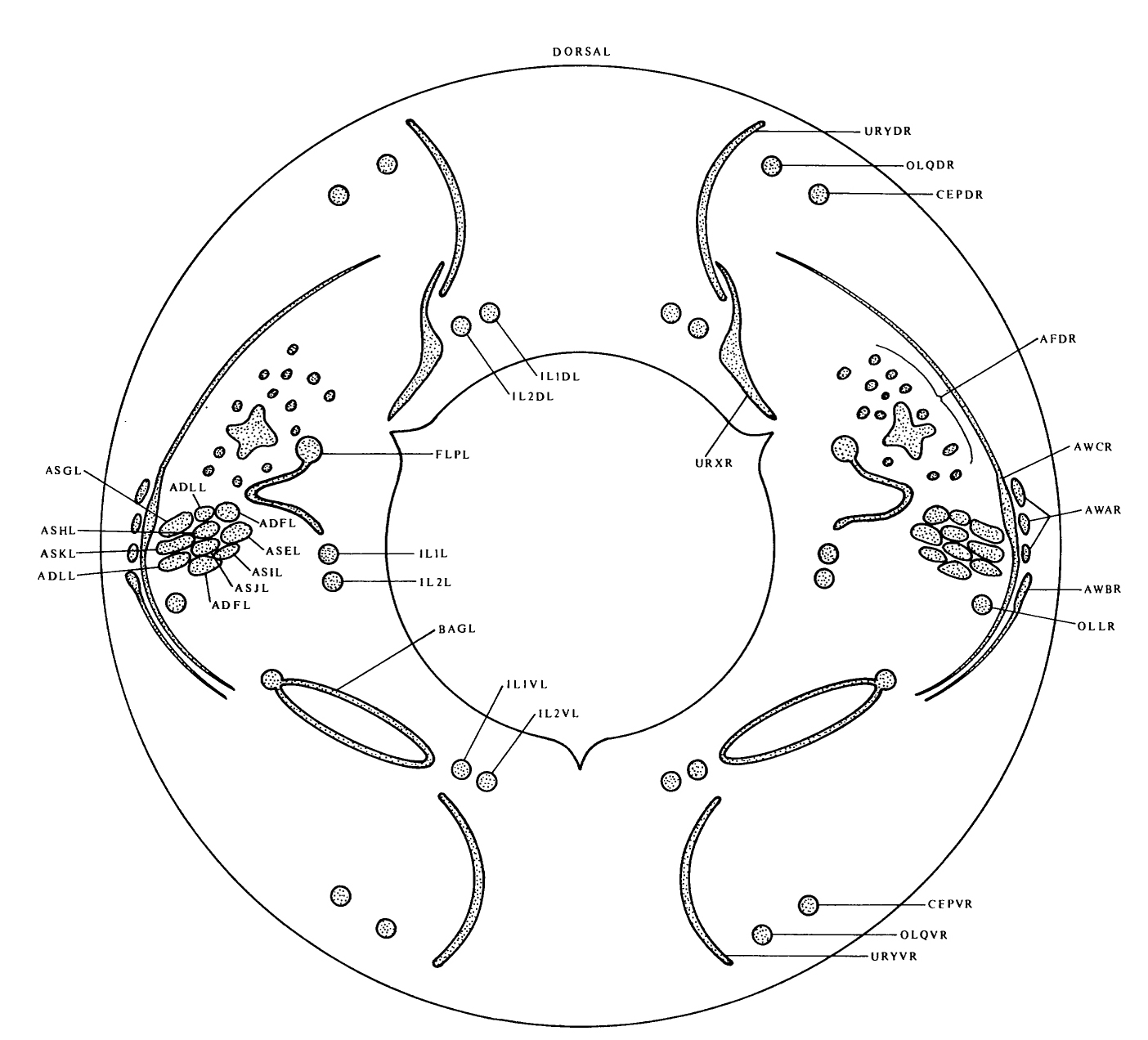

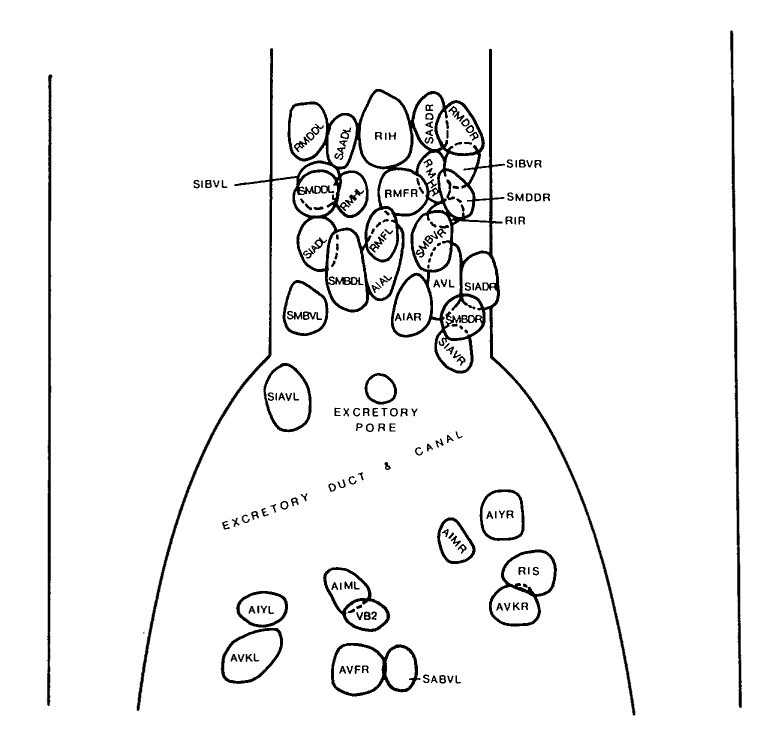
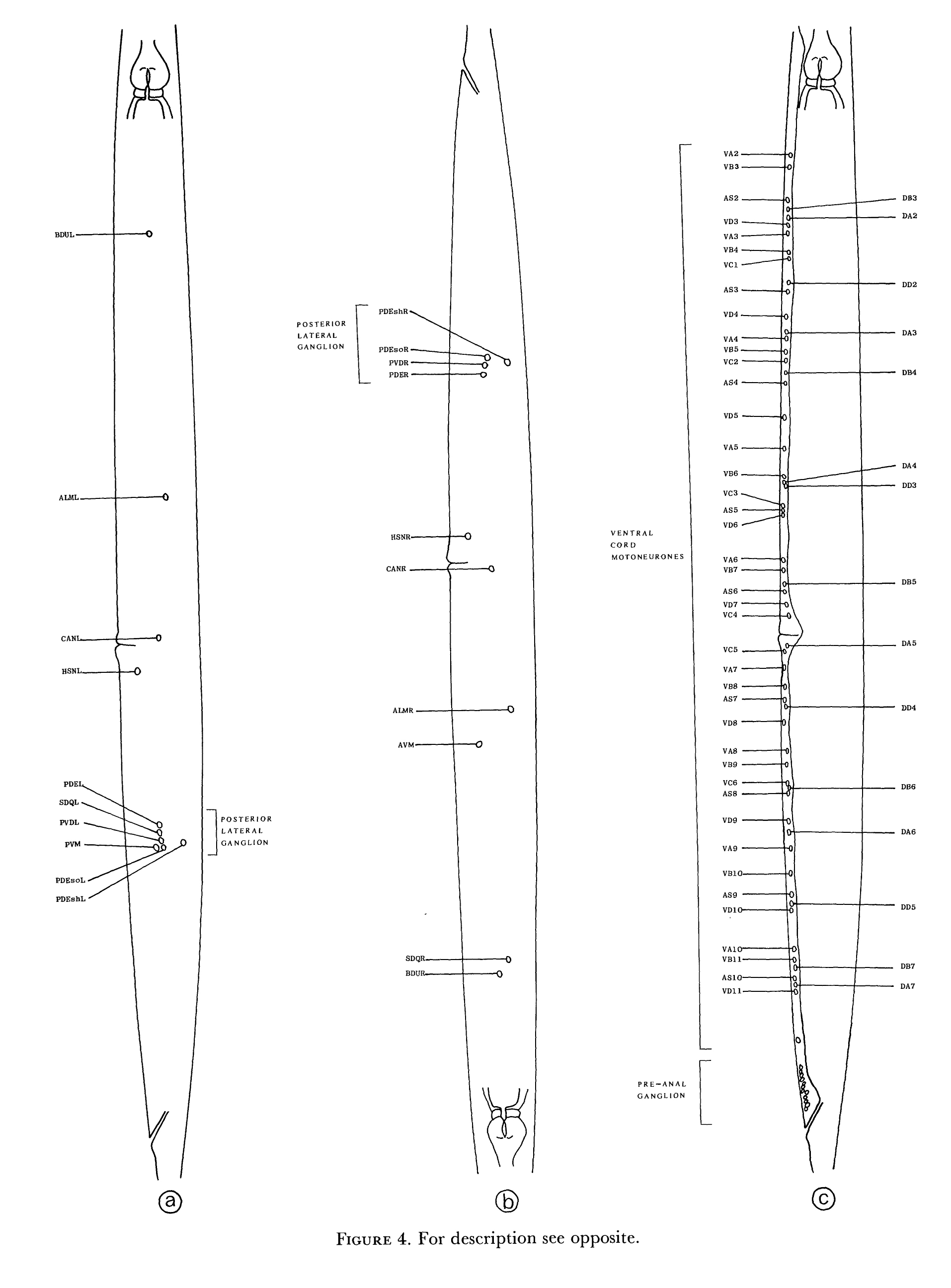



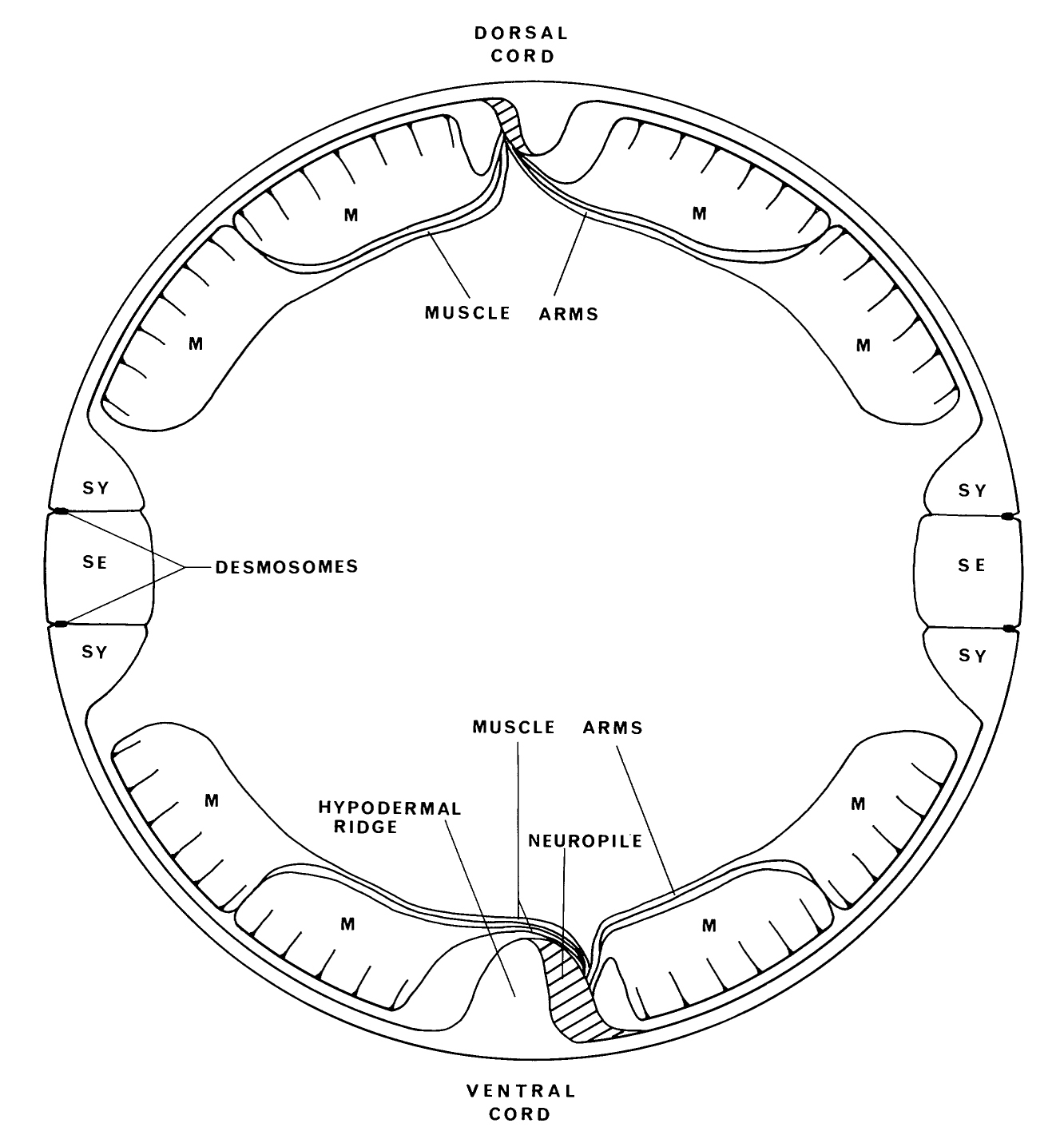
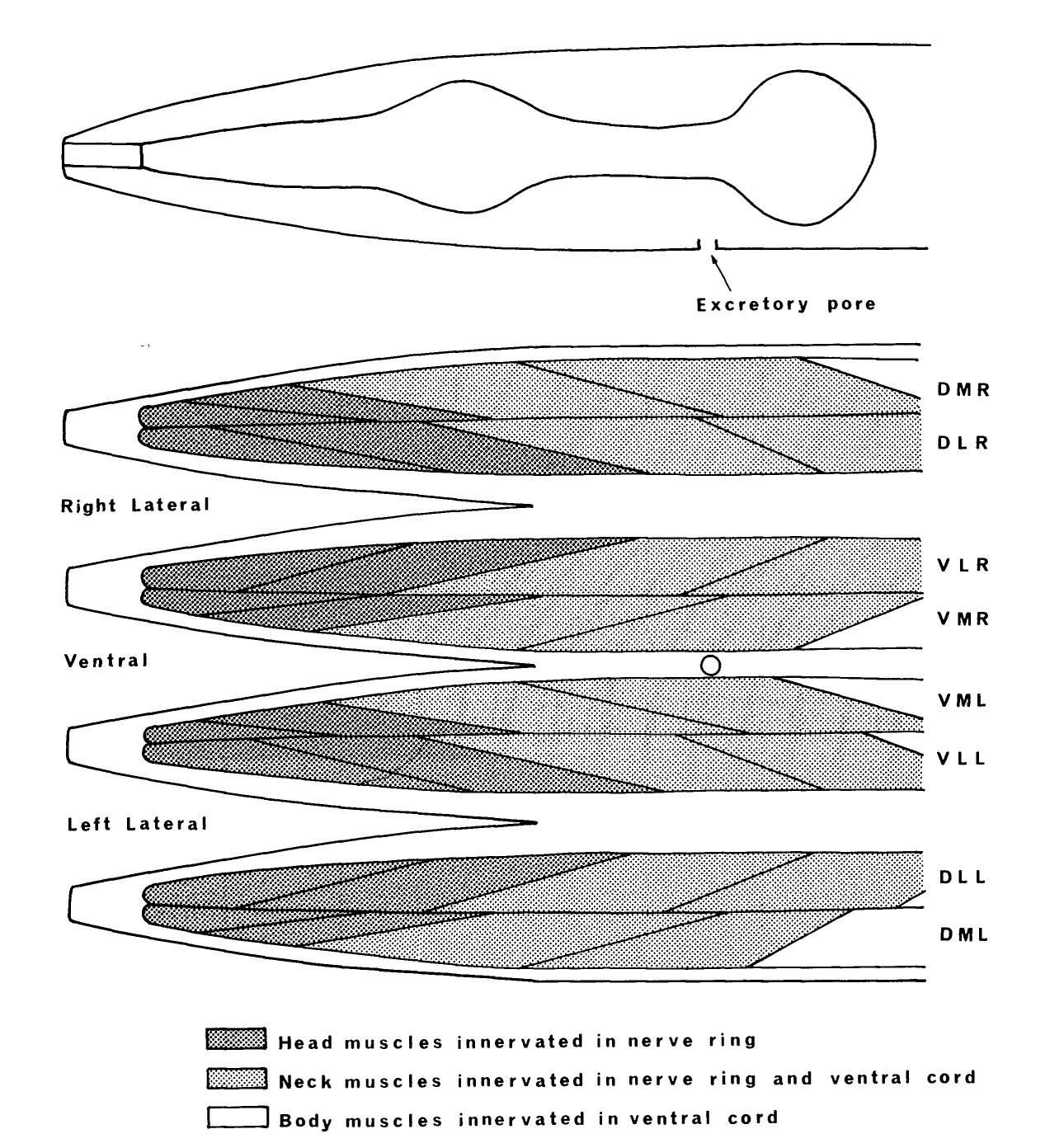


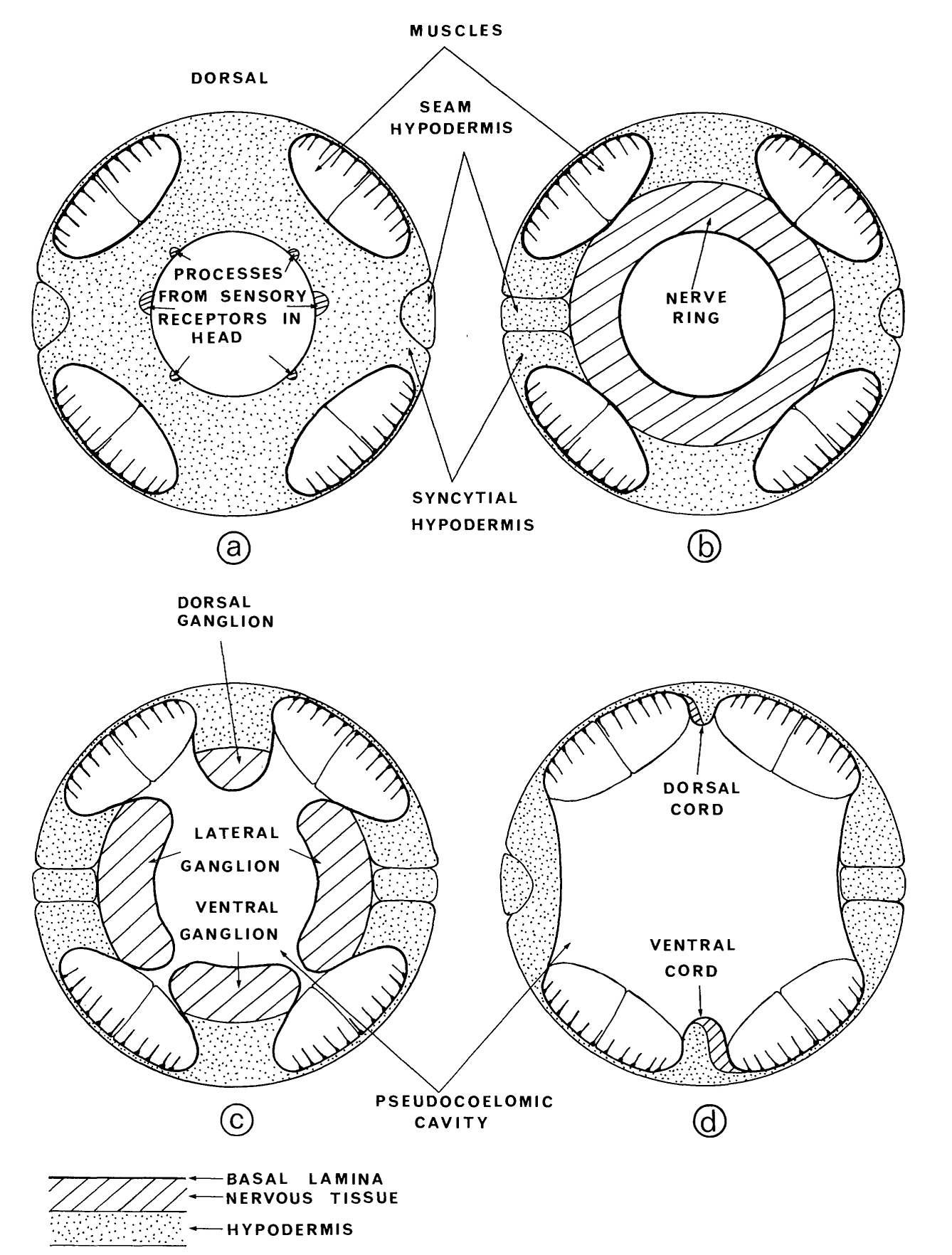
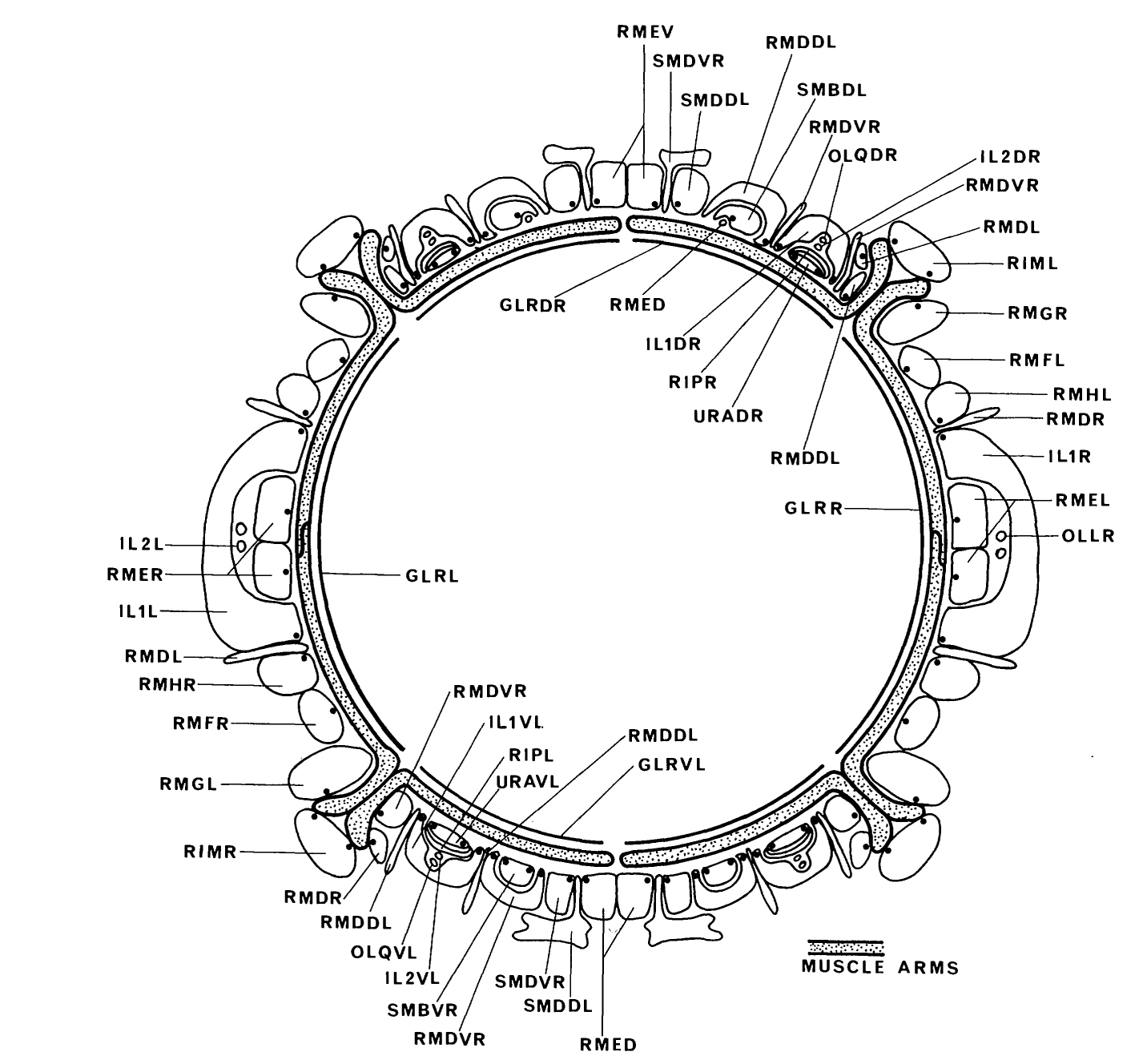
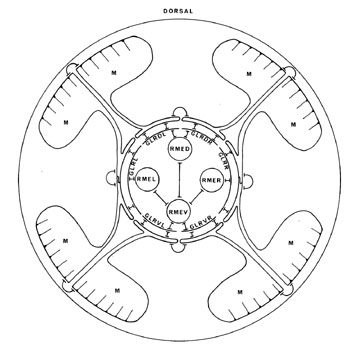
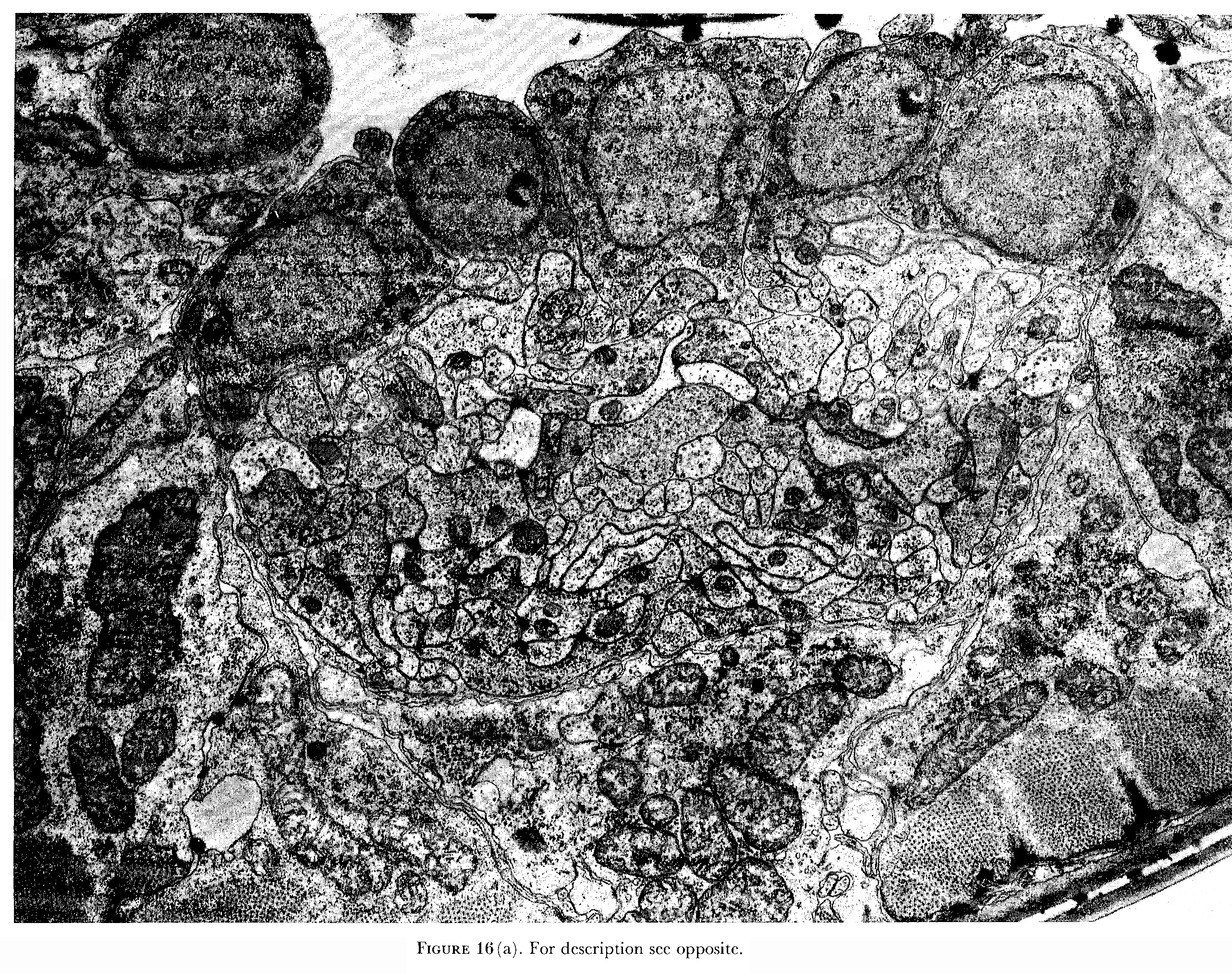
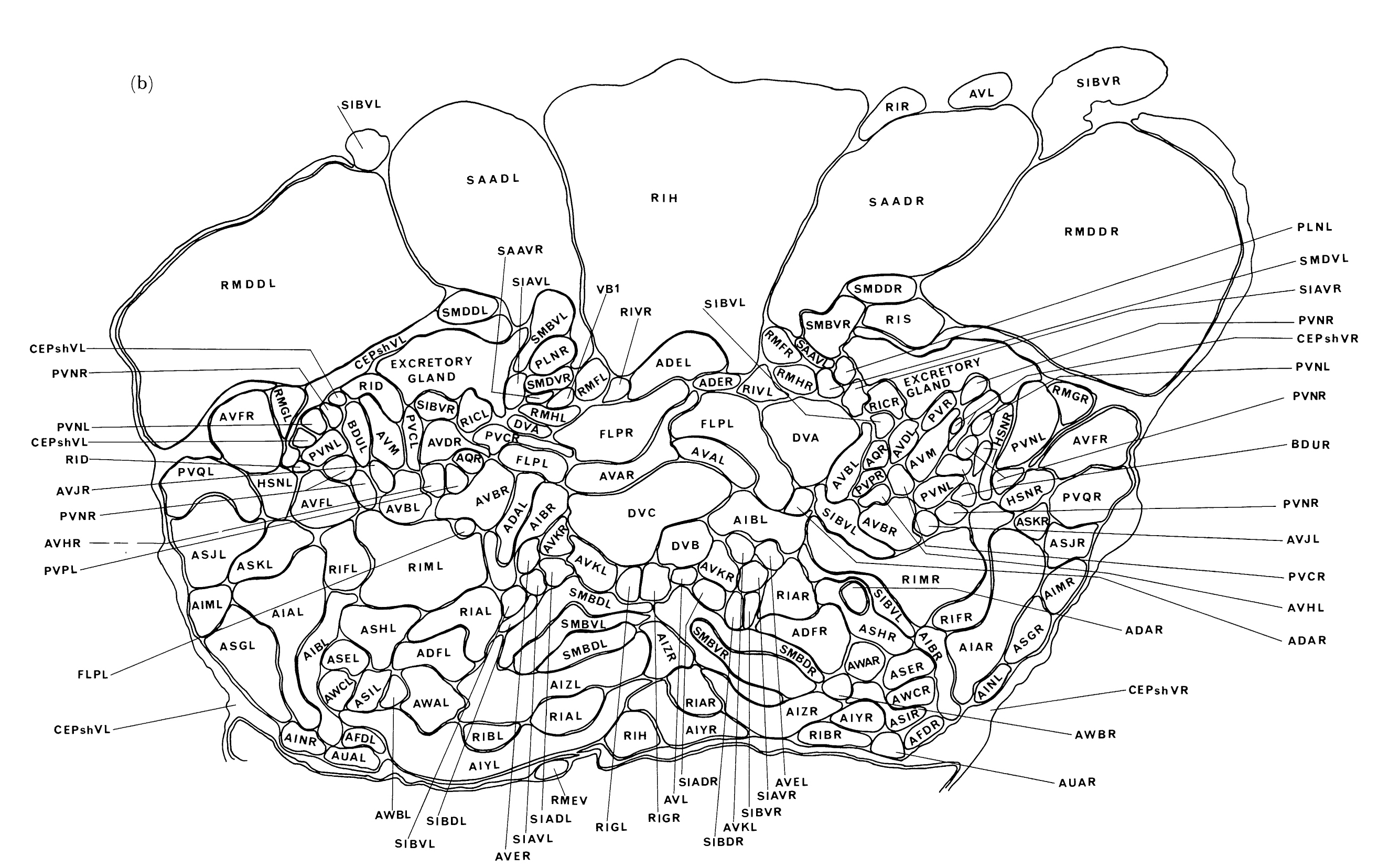
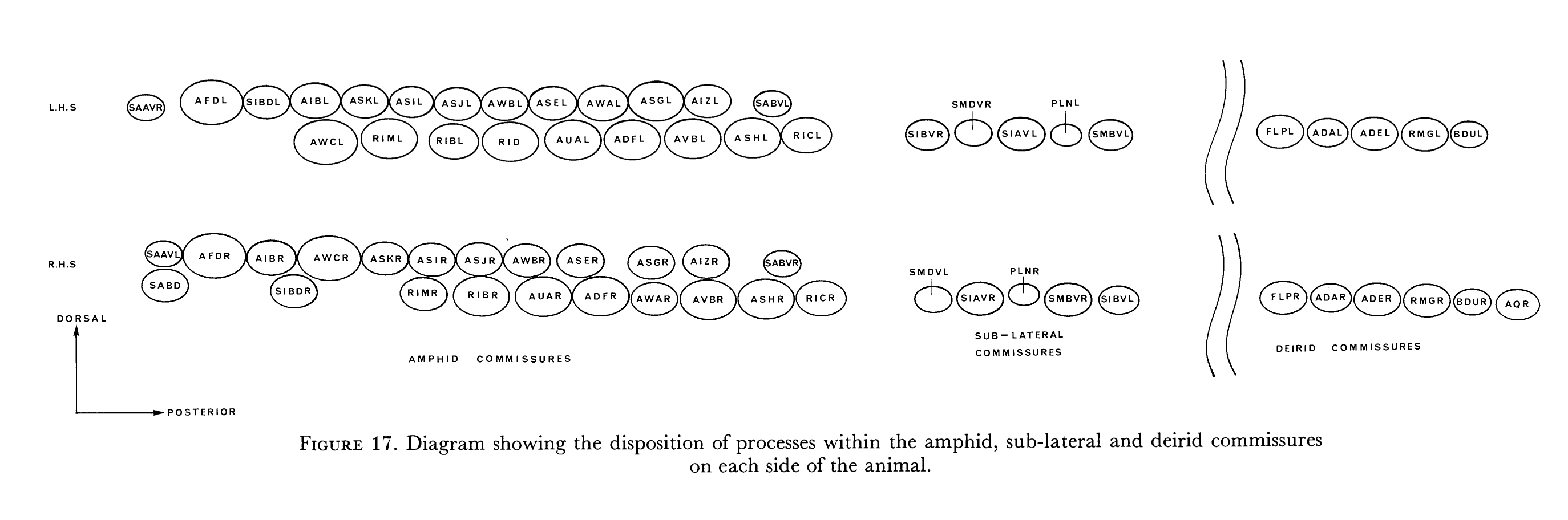
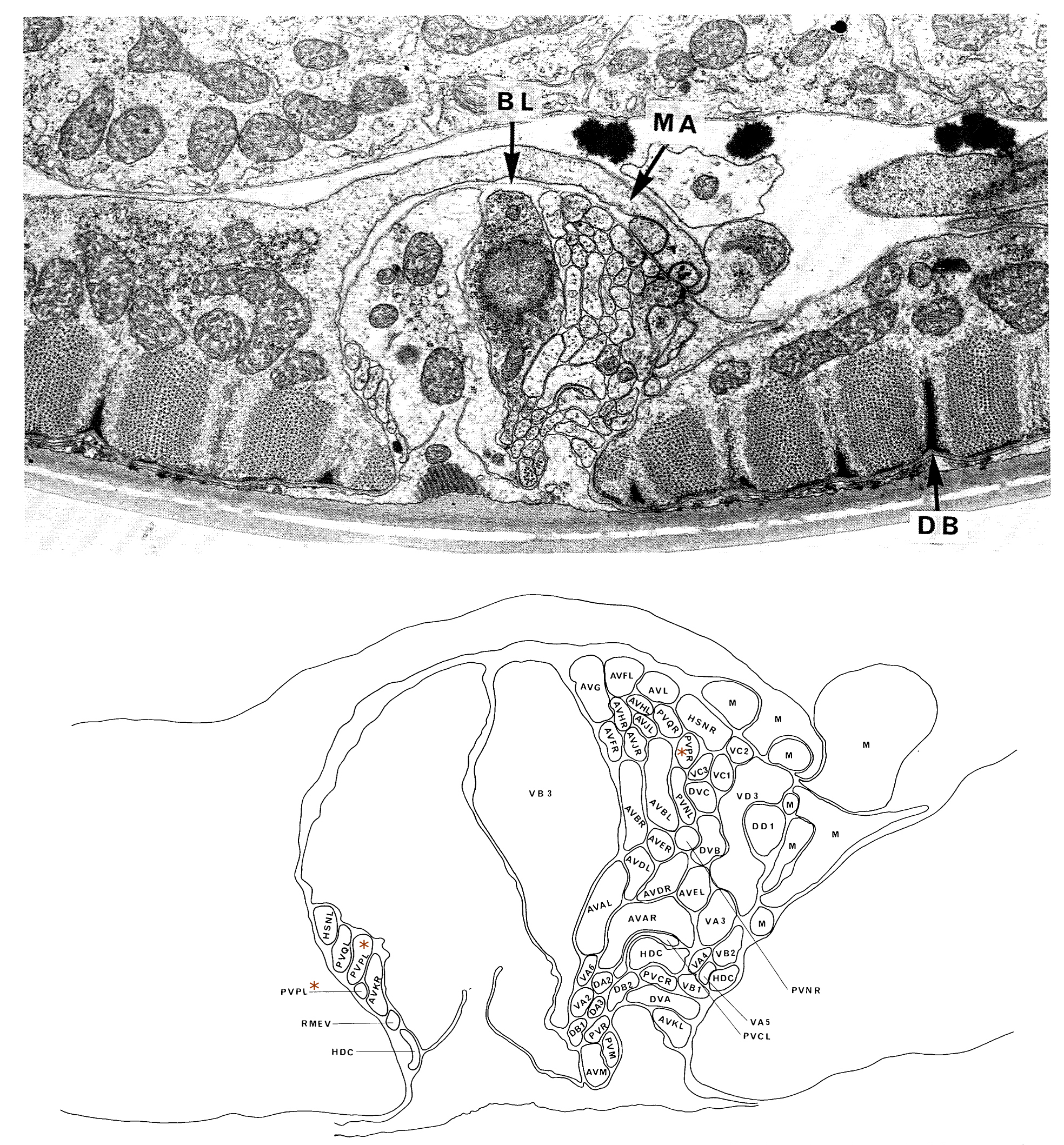
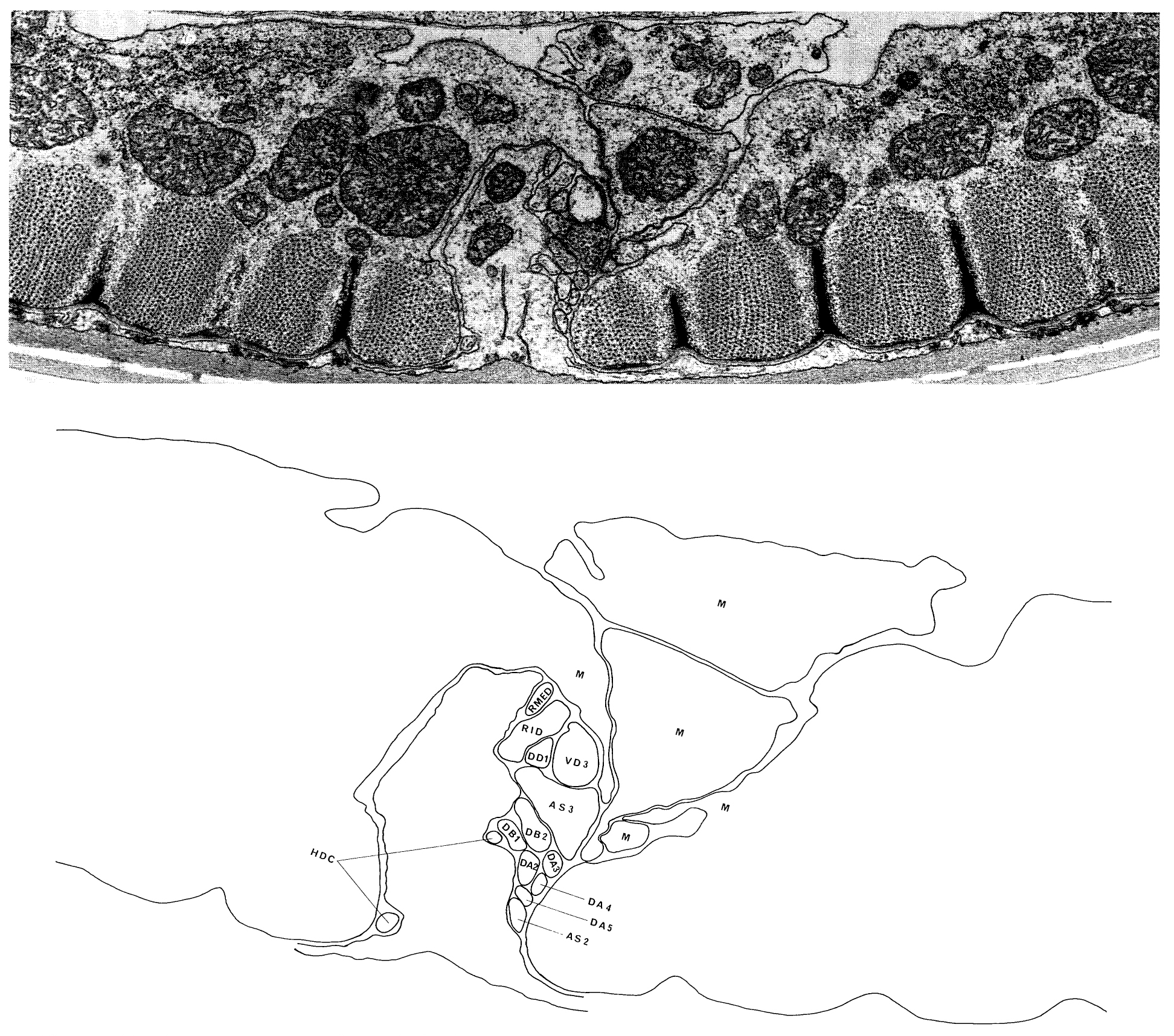
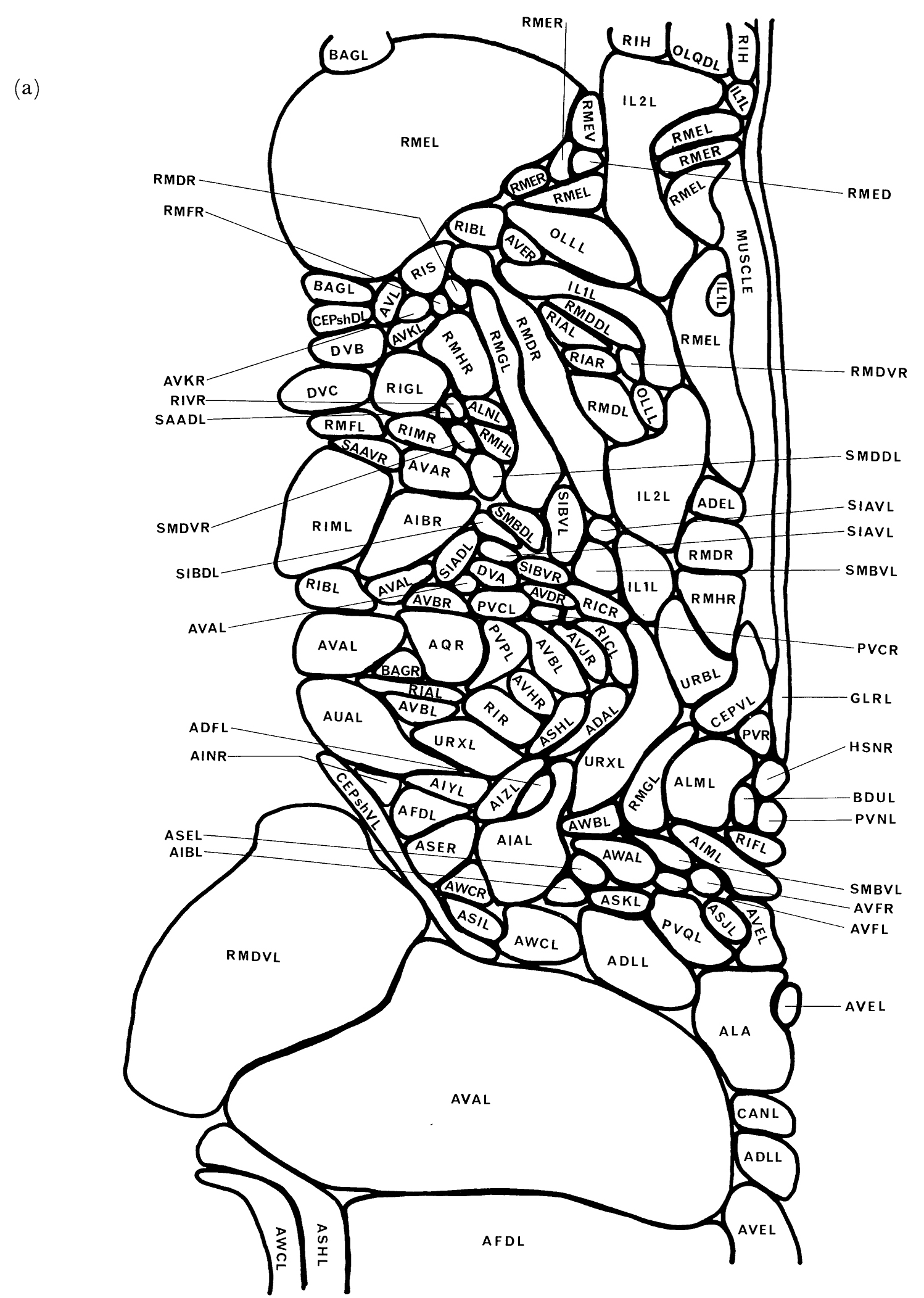

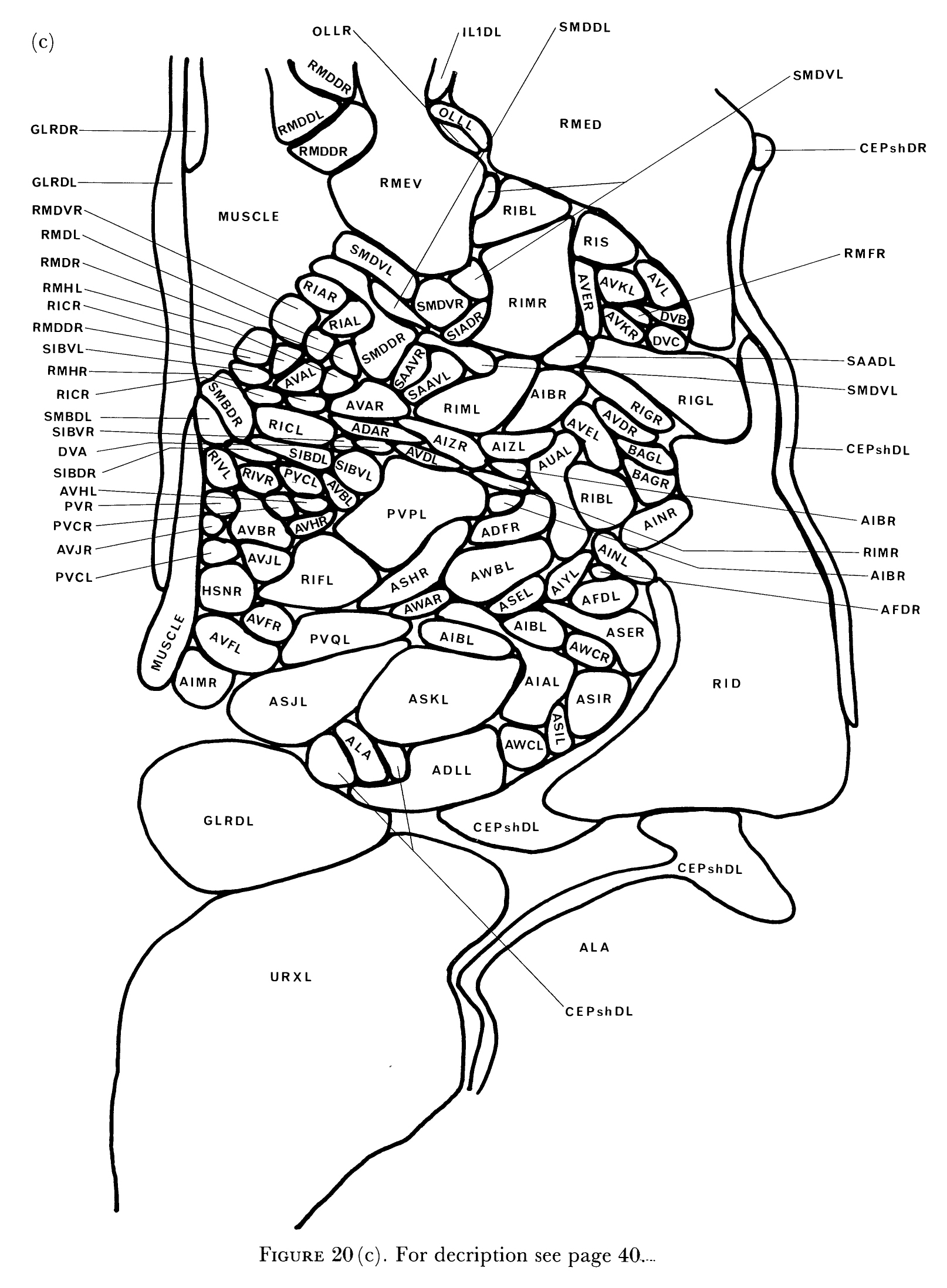
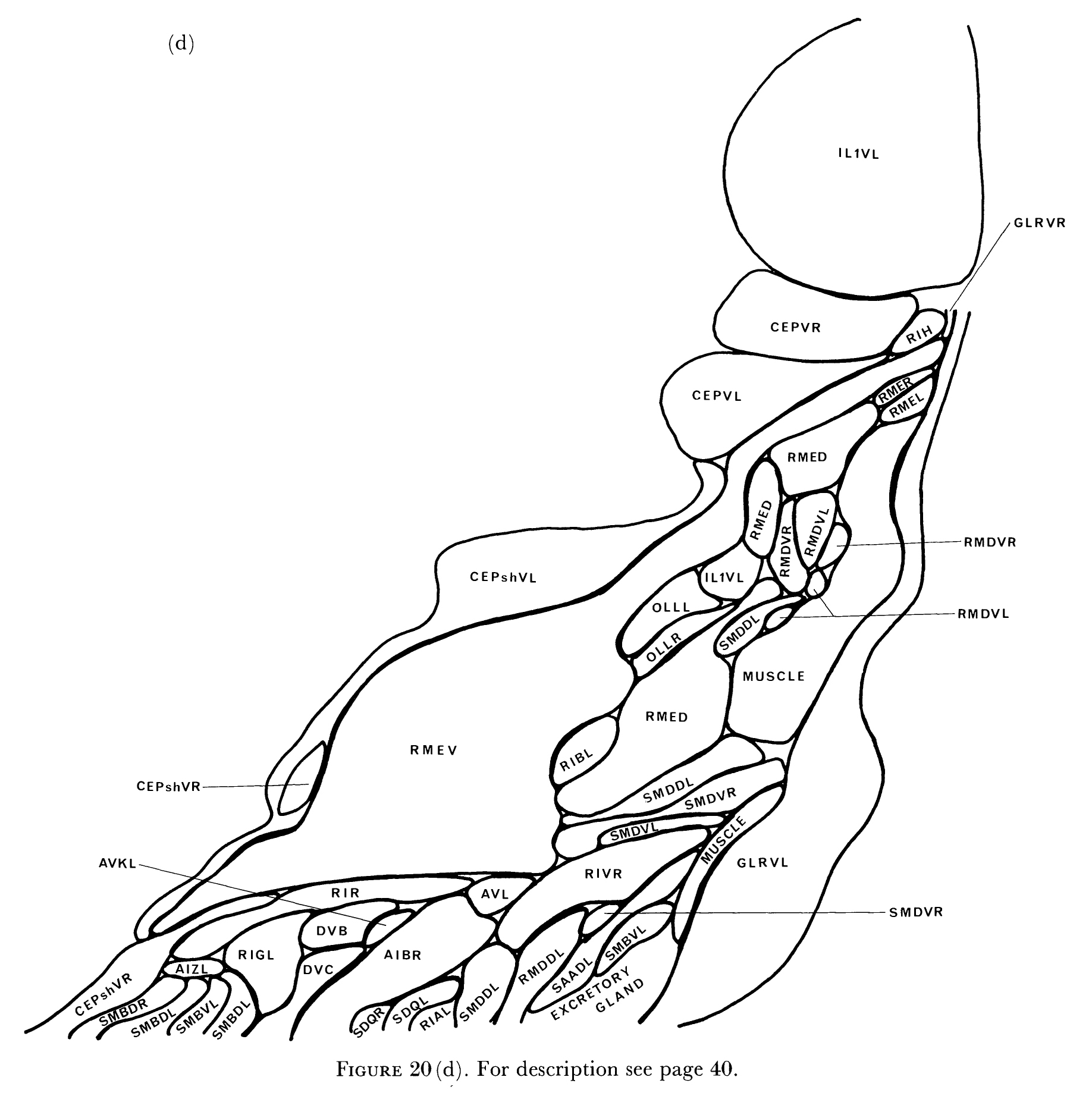
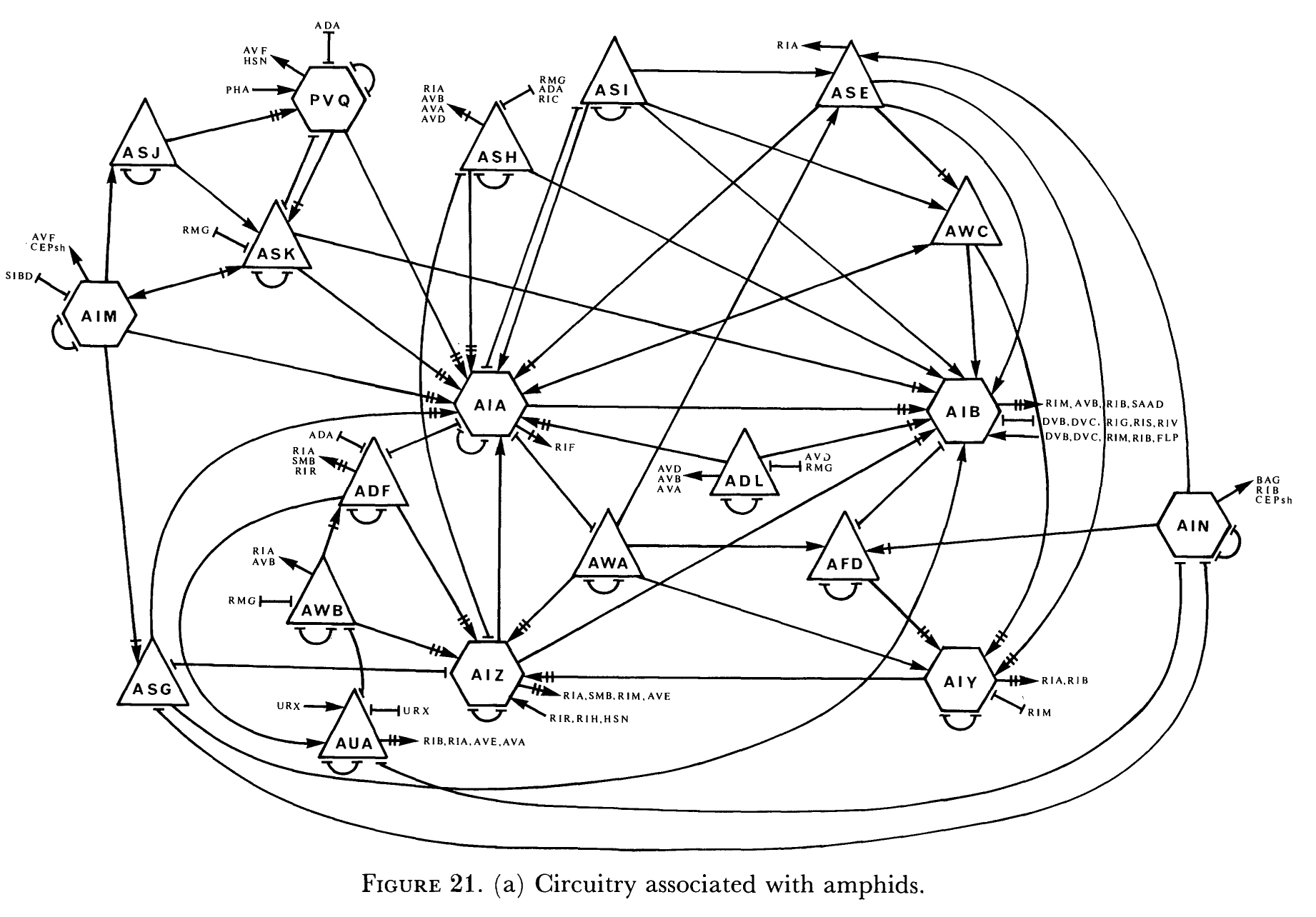
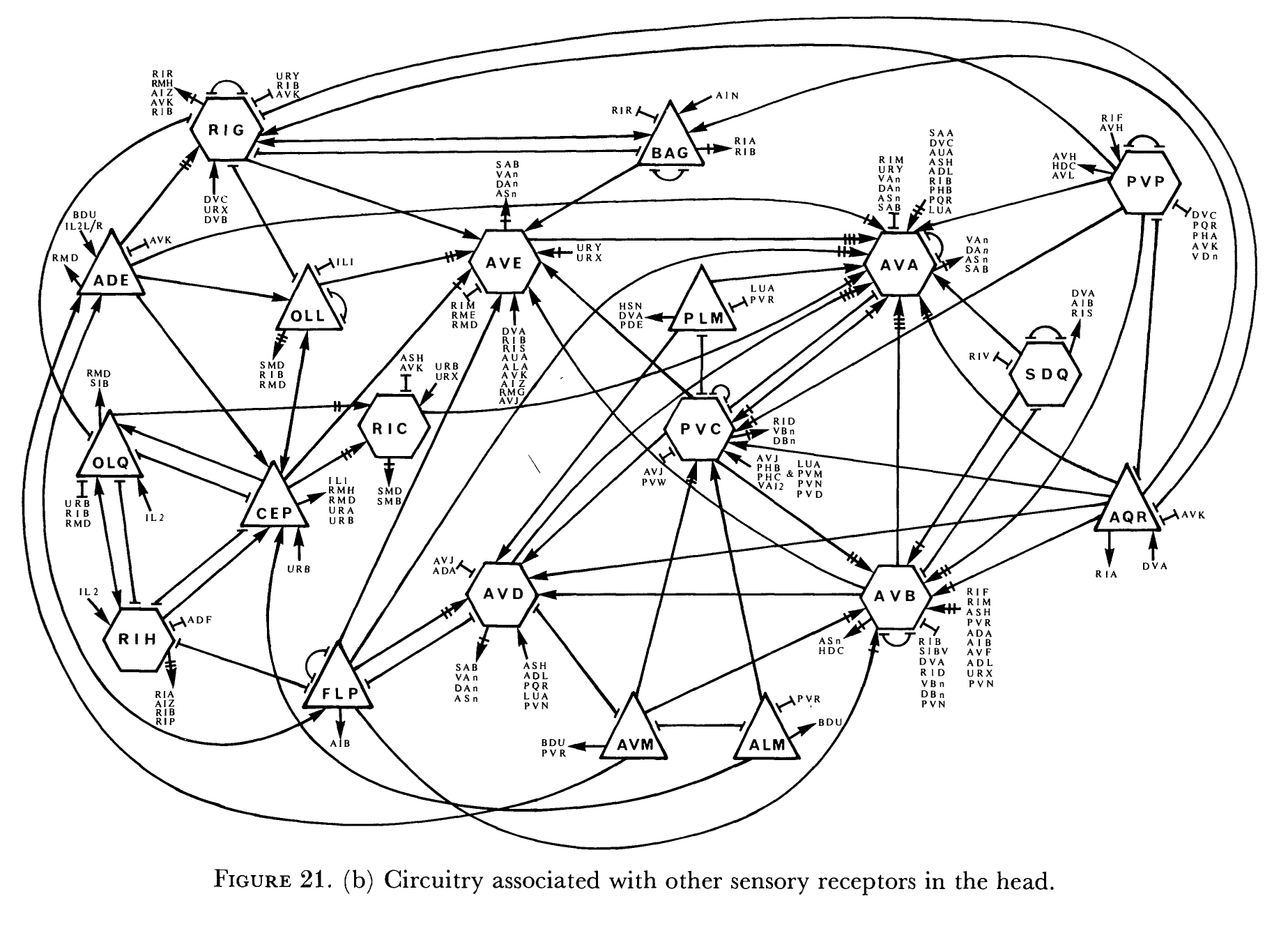

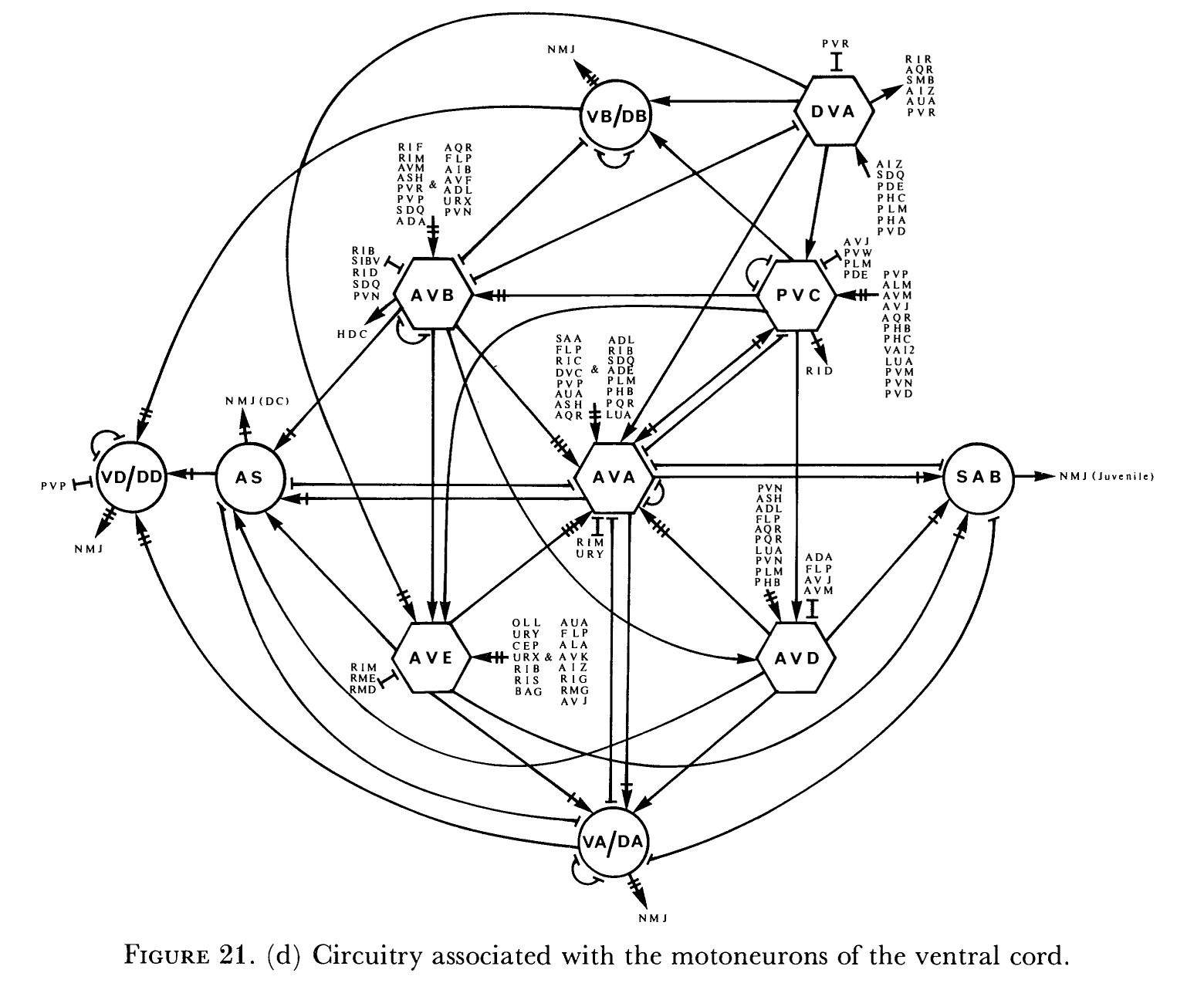

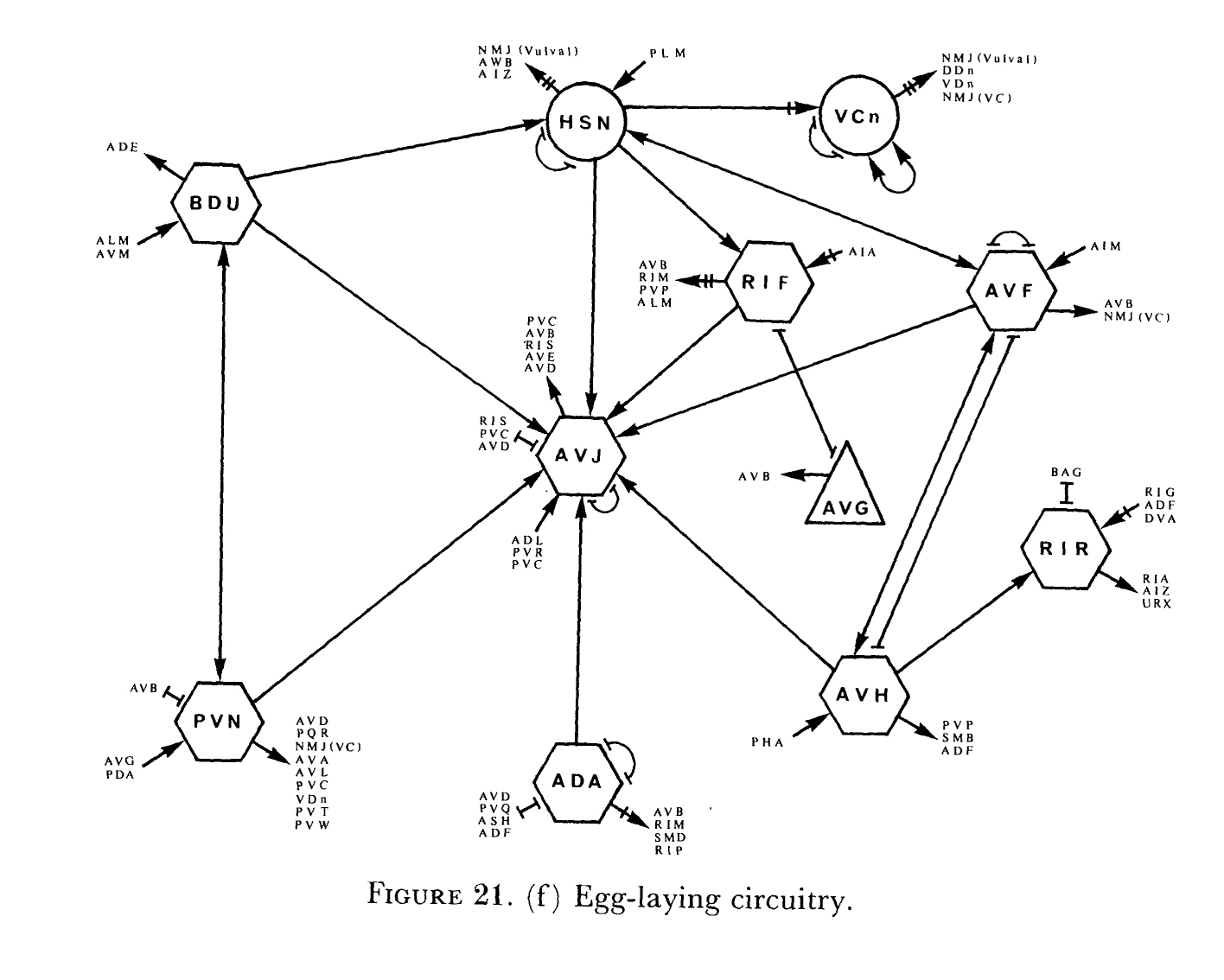
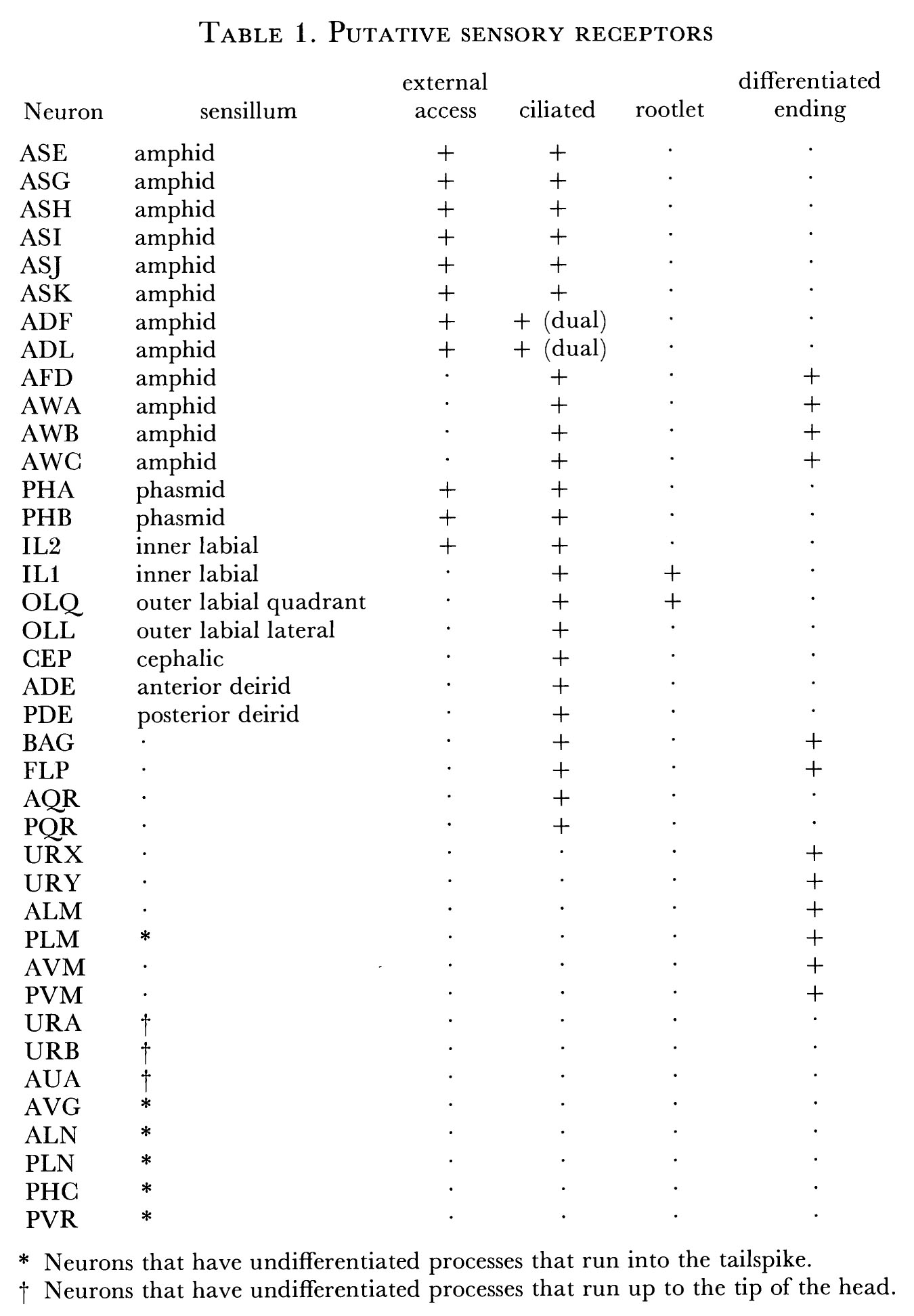

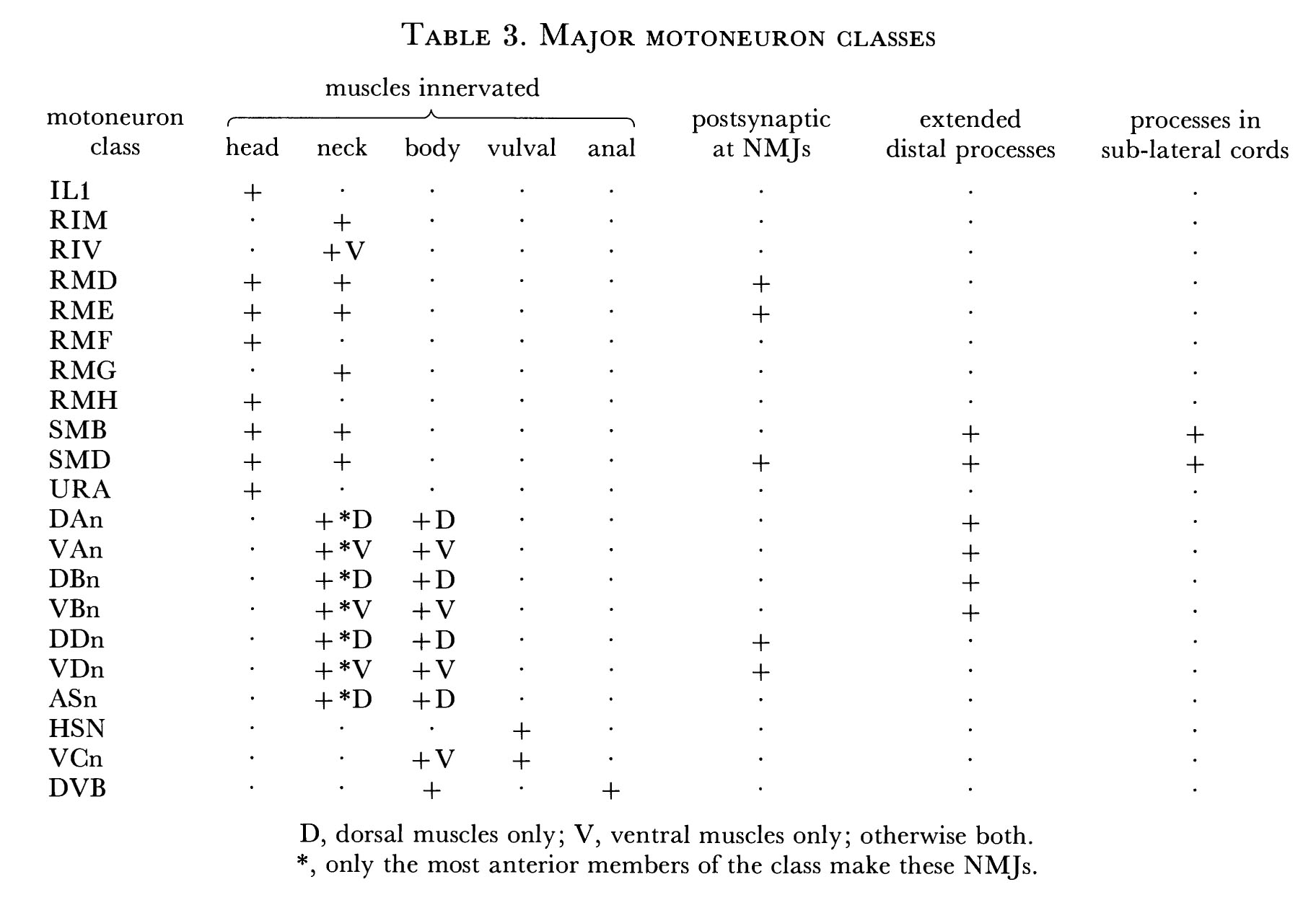

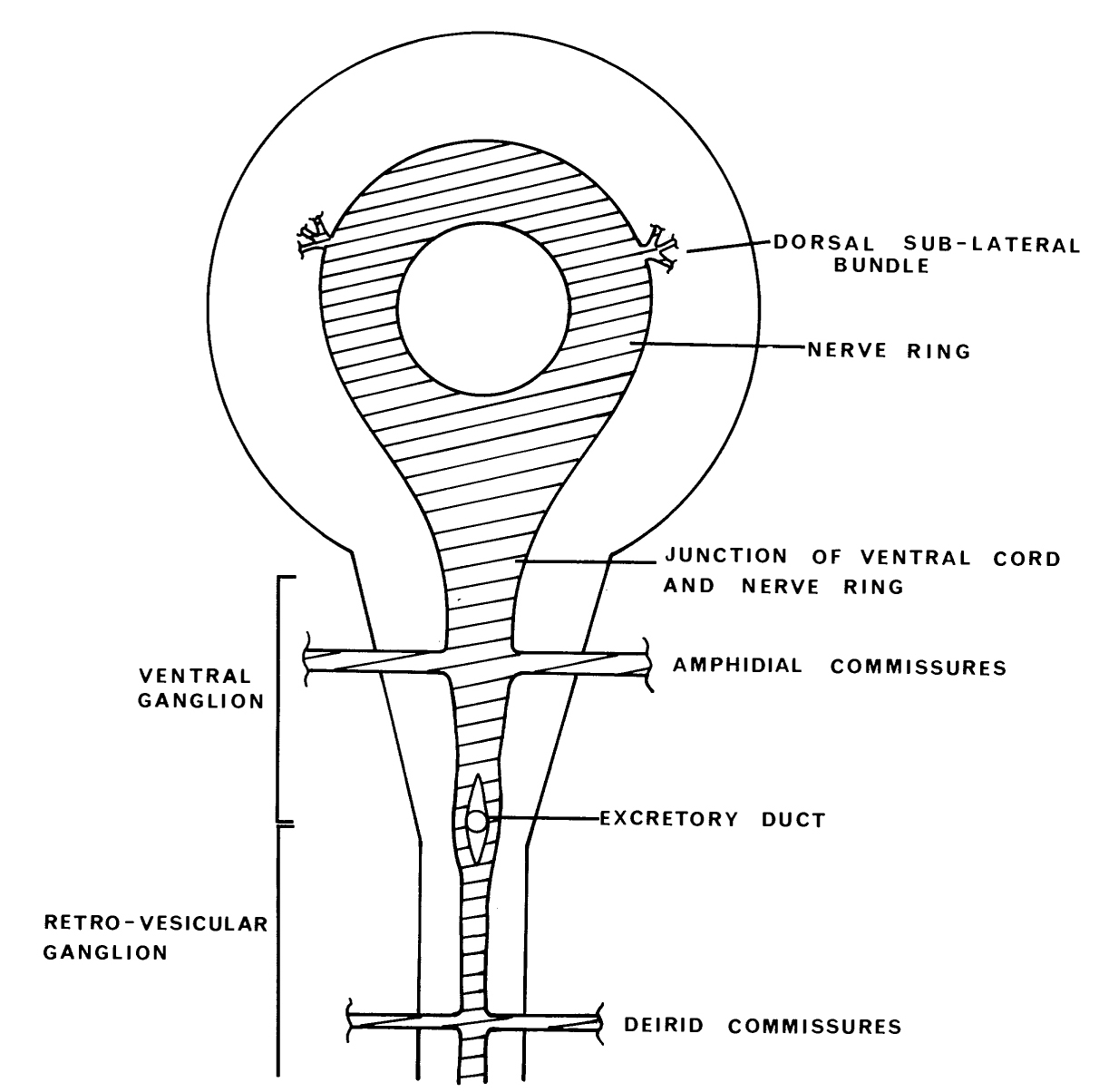
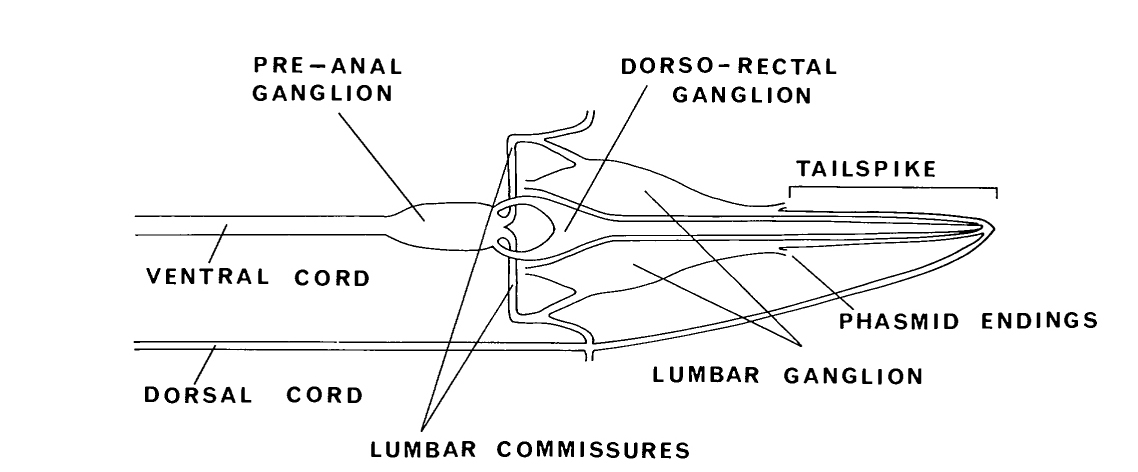
 Click picture for higher resolution image
Click picture for higher resolution image

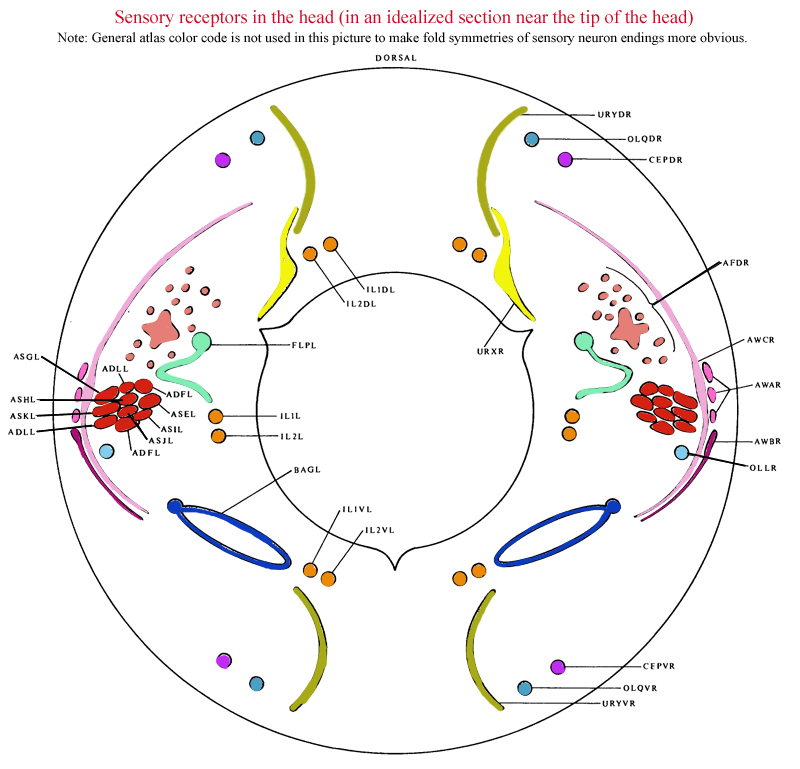{kind=link}
{kind=link}
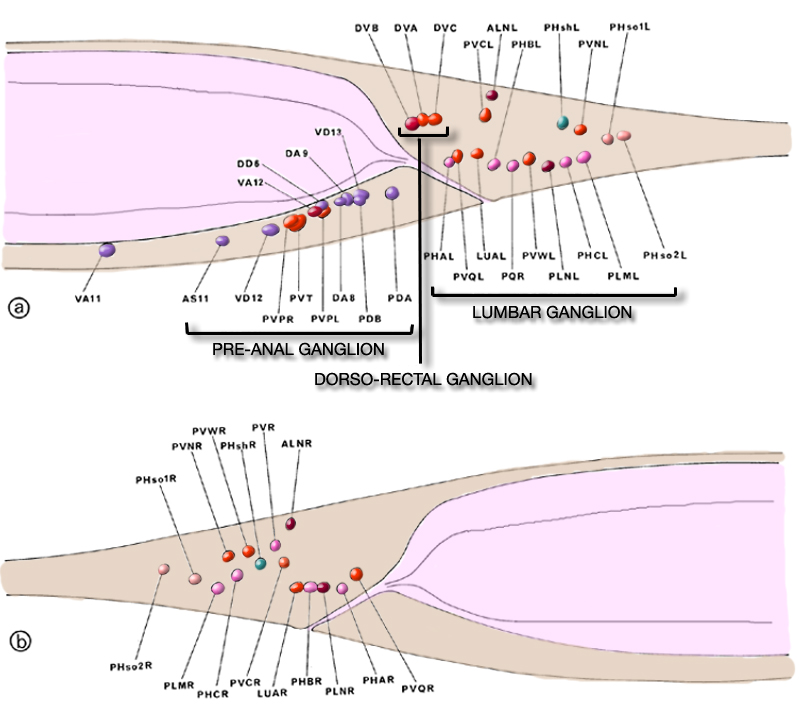{kind=link}
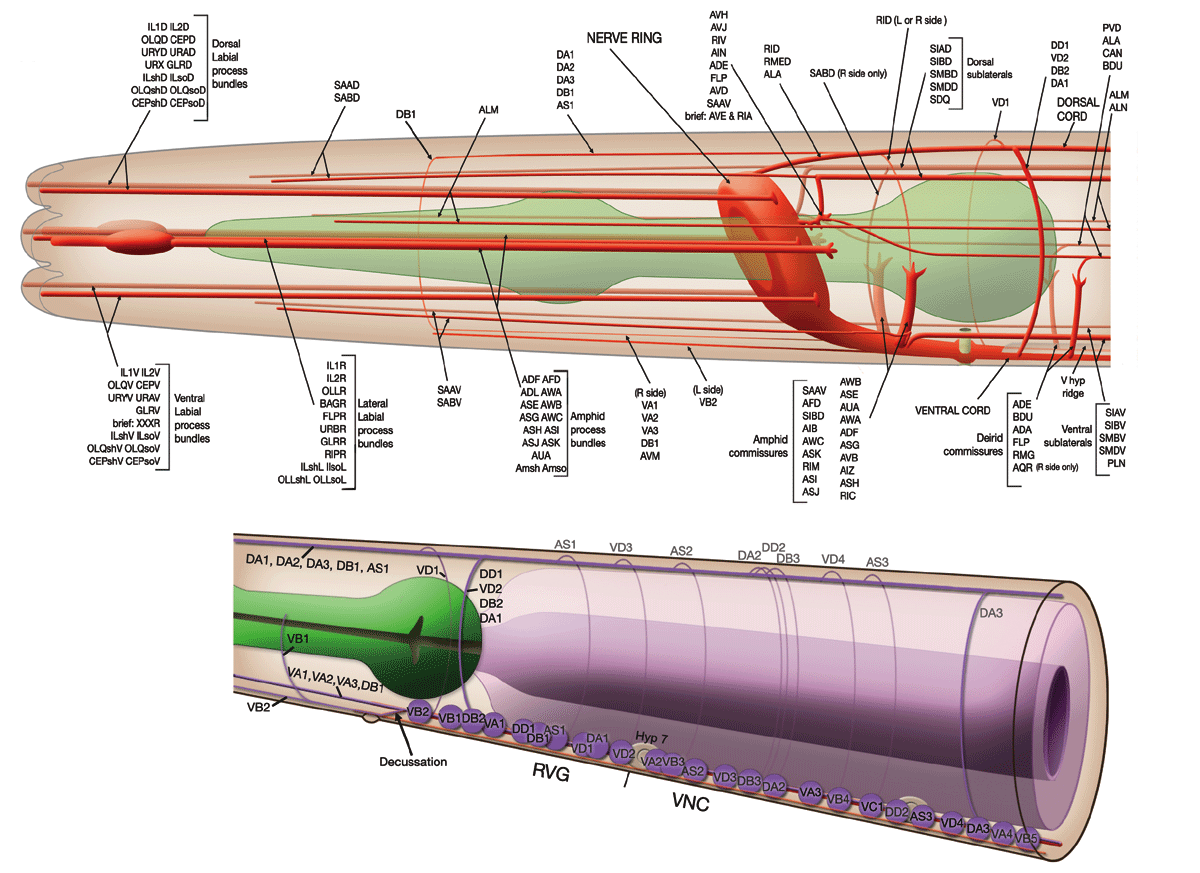{kind=link}